| 京都大学野生動物研究センター > 屋久島フィールドワーク講座>第10回・2008年の活動−ヤモリ班−報告書 |
| 概要 | サル班 | シカ班 | ヤモリ班 | 博物館班 | シンポジウム | スタッフ |
| 報告書 | 感想文 |
講師・チューター:疋田努・戸田守・河合潮
受講生:小島光明・坂田ゆず・前田真希・正井佐知
九州南部から大隅諸島にかけての地域には形態的に類似した2種のヤモリ(ヤクヤモリGekko yakuensisとミナミヤモリG. hokouensis)が分布する。これら2種は九州南部の広い範囲で同所的にみられるが、大隅諸島ではほぼ異所的に分布し、多くの島嶼ではどちらか片方の種のみしかみられない(竹内・戸田,2006)。このように、面積が限られた島嶼において近縁な2種が排他的に分布する状況は、これらの種が競争関係にあることを示唆している。そのなかで、屋久島は島嶼部としては例外的にヤクヤモリとミナミヤモリの双方が分布する島となっており、両種の分布の成因や種間関係などを論じるうえで興味深い。
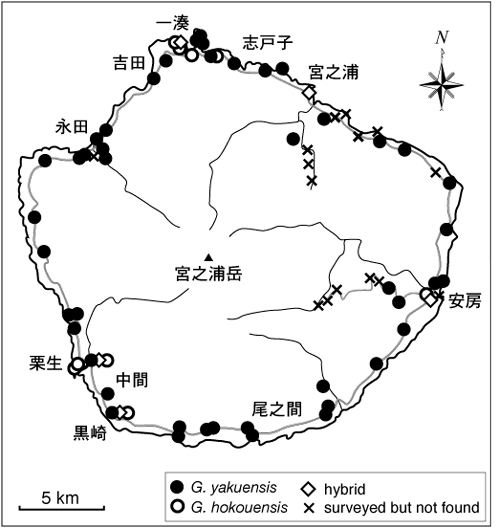
このような背景のもと、2005年度と2006年度の屋久島フィールドワーク講座では屋久島内における2種のヤモリの分布調査がなされ、ヤクヤモリが島内の低地全域に広く分布するのに対し、ミナミヤモリは一湊、宮之浦、安房、栗生など一部の集落に局所的に分布することが明らかになった(図1)。ミナミヤモリが分布するこれらの集落は、いずれも現在、あるいは過去に島外との交易に使われていた港を擁し、逆にそのような港がない集落にはミナミヤモリが分布しないことから、屋久島のミナミヤモリは物資の搬入に伴う人為的な持ち込みに由来する可能性が強く示唆される。
もし、屋久島のミナミヤモリが人為移入に由来するならば、その侵入によって、在来種であるヤクヤモリが様々な影響を受けていると予想される。特に、これらのヤモリは九州の一部の地域で交雑集団を形成していることが知られているため、屋久島でも種間交雑が進行し、ヤクヤモリ集団の遺伝的構成が変化してしまう可能性も考えられる。交雑による遺伝的構成の変化は、その種が進化の過程で獲得してきた地域適応的な特徴を崩壊させてしまう場合もある。したがって、屋久島におけるミナミヤモリの分布や種間交雑の進行状況について正確に把握することは、生物多様性保全の観点からも重要である。上述したように、屋久島におけるミナミヤモリの分布は大枠で分かっているものの、ミナミヤモリが定着している地域においてこの種がどのように分布拡大を果たしているかなど、その詳細まで十分に分かっているとは言い難い。そこで今年度は、屋久島においてミナミヤモリが比較的多く見つかっている栗生周辺と一湊周辺に狙いを絞って、より細かい地理スケールで2種のヤモリの分布を調べ、さらに地理情報システムを用いて、ミナミヤモリや交雑個体の分布と環境との関係について分析した。
図1.2005年度および2006年度の調査で得られたヤモリ類の分布記録
野外調査は2008年8月18日から同年8月22日にかけて栗生周辺と一湊周辺で実施した。事前に、環境省自然環境局生物多様性センターが運営する生物多性情報システム(http://www.biodic.go.jp/J-IBIS.html)を基に土地利用区分を示した地図を準備し、それを参考にして、なるべく様々な環境を網羅するよう調査区域を設定した。各調査区域では、主に夜間にヤモリの好みそうな場所(電柱、樹木、公衆トイレ、学校の校舎、神社、道路沿いののり面、架橋の欄干など)を探索し、ヤモリの発見に努めた。また、補助的に行った日中の調査ではヤモリ類が潜んでいそうな隙間を覗いて休止中の個体の発見に努めた。
ヤモリが発見された場合はそのまま手で押さえるか、釣竿で追い落としてから捕獲した。また、隙間に潜んでいるヤモリは細い棒などで追い出して捕獲した。捕獲時にはできるだけ胴体を押さえつけるようにし、尾を自切しないよう注意を払った。
捕獲したヤモリは1個体ずつビニール袋に入れ、通し番号をふってから、発見者、発見時刻、天気、地名、緯度経度、周囲の環境、底質を記録し、さらにヤモリの外部形態形質を観察して、種の同定および性別の判定を行った。形態的特徴から確実に種の同定ができた個体は、原則として、記録終了後に元の場所に放逐した。放逐の際は市販のマジックで腹部に一時的な標識を施し、未記録の個体と区別がつくようにした。一方、形態的特徴から交雑個体と判定されたものや、分布の状況から潜在的に交雑の可能性が疑われる地点の個体はそのまま研修センターに持ち帰り、標本にした。緯度経度の測定には全地球測位システム(GPS V: GARMIN社)を使用した。
ヤモリの同定は、先行研究に従い、間鼻板のサイズと尾の大型鱗の状態によって行った(表1)。ただし、尾の大型鱗は、尾が根本付近から欠損した個体や再生尾である個体ではその状態を決定できないため、そのような個体については間鼻板のサイズのみを観察した。とはいえ、間鼻板のサイズには各々の種のなかにもかなりの変異が見られるうえ(岡田,1998; Toda et al., 2001)、生体でその状態を正確に評価するのがやや困難であることから、この形質のみによって種を同定するのは危険である。そこで、間鼻板のサイズのみによる同定は暫定的なものとし、それらの個体の記録は環境の分析から除外した。
交雑個体は、尾の大型鱗の状態と間鼻板サイズをともに決定できた個体のうち、2形質の状態が互いに矛盾するものをそれとみなした(表1)。ヤクヤモリとミナミヤモリは野外でも雑種第2代以降の個体や戻し交雑個体を生じ、そのような個体は、形態形質のみの観察ではしばしば純系親種との識別が困難である(Toda et al., 2001)。つまり、上記の同定基準では交雑個体の出現が過小評価される傾向がある。この問題を解消するためには、分子形質を用いた分析を行う必要があるが、本調査期間のなかでは、後に予定しているそのような分析のための凍結組織サンプルを採取するに留めざるを得なかった。とはいえ、ここで採用した基準で交雑個体を特定することにより、少なくともその記録地点では2種の交雑が起きていると見なすことができる。
表1. 形態形質に基づくヤクヤモリとミナミヤモリの識別点
| ヤクヤモリ | ミナミヤモリ | |
|---|---|---|
| 間鼻板サイズ1) | その長さは後続の鱗の1.5倍を越える2), 3) | 後続の鱗の1.2倍を超えない3) |
| 尾の大型鱗4) | 少なくとも,基部から第13自切溝 まで続く | 無いか,あっても第11自切溝より先端へは続かない |
組織サンプルの摘出と標本の作製は次の手順で行った。まず、麻酔剤を用いて採集したヤモリを安楽死させた後、個体ごとにラベルを着けた。次に、解剖バサミを用いて腹部と大腿部を切開し、各個体から肝臓片と筋肉片を摘出した。取り出した組織片は専用のサンプルチューブに入れて液体窒素中で凍結・保存した。最後に、プラスチック容器の底に10%ホルマリンを浸した紙を敷き、その上で組織片摘出後の個体の形を整え、固定した。なお、これらの標本はすべて京都大学総合博物館の爬虫類コレクションに登録した。
ヤクヤモリとミナミヤモリ、および交雑個体の出現環境を比較するため、次の2通りの解析を行った。 それは、1)ヤモリの記録地点の属性検索による解析と、2)バッファ解析である。1の解析では、地理情報システムを用いて、各個体が記録された地点の土地利用タイプを検索し、それを種ごとに集計して比較した。土地利用タイプは、便宜的に、a) 市街地、b) 農耕地(樹園地を含む)、c) 森林(自然林、二次林、植林を含む)、d) 草原・自然裸地の4カテゴリーにまとめた。 一方、2の解析では、まず各ヤモリの記録地点を中心とする半径100 mの区域(バッファ・ゾーン)を地図から切り出し、次に、その区域のなかに占める森林面積の割合(%)を「森林度」として求め、ぞれぞれの種がどのような森林度で出現するかを評価した。すなわち、1の解析ではヤモリの記録地点そのものの環境を、2の解析ではヤモリの記録地点の周囲の環境を評価したことになる。これらの解析は、ともに地理情報システム解析ソフト(Arc GIS, ver 9.2)を用いて行った。
野外調査の結果、栗生周辺で62個体、一湊周辺で49個体(合計111個体)のヤモリが発見・捕獲された。このうち、栗生周辺の44個体と一湊周辺の34個体は少なくとも尾の先端付近まで非再生尾であったため、尾の大型鱗の状態と間鼻板サイズの2形質に基づいて確実に同定できた(表2)。一方、一湊周辺で捕獲された2個体は尾が基部付近から再生尾であり、さらに間鼻板も中間的なサイズであったため種の帰属が決定できなかった(表2)。
表2.栗生周辺と一湊周辺で記録されたヤモリの個体数。括弧内は間鼻板のサイズのみによって暫定的に同定された個体を含む数字
| ヤクヤモリ | ミナミヤモリ | 交雑個体 | 同定保留 | 合計 | |
|---|---|---|---|---|---|
| 栗生周辺 | 23 (34) | 17 (23) | 4 (5) | 44 (62) | |
| 一湊周辺 | 17 (26) | 16 (20) | 1 | (2) | 34 (49) |
これらの記録を、地理情報システムを利用して作製した土地利用図の上にプロットしたのが図2である。また図3には、2形質を用いて確実に同定された78個体のデータについて、記録地点の土地利用タイプの内訳を種ごとに示した。これらの図にみるように、ミナミヤモリが主に市街地と農耕地で記録されたのに対し、ヤクヤモリはそのほとんどが森林で記録された。また、交雑個体は市街地・農耕地と森林の双方で記録された。
次に、バッファ解析の結果を図4に示す。ヤクヤモリのほんどは、周囲を完全に森林に囲まれた環境(森林度100 %)で記録されたが、少数個体が森林度30-40 %の環境でも記録された。一方、ミナミヤモリは、森林度が低めの環境(10-40 %)で、より頻繁に記録される傾向が認められたが、森林度が高い環境(50-100 %)からもかなりの個体が記録され、全体としてばらつきが非常に大きかった。交雑個体の記録地点の森林度は30-100 %の範囲であった(図4)。
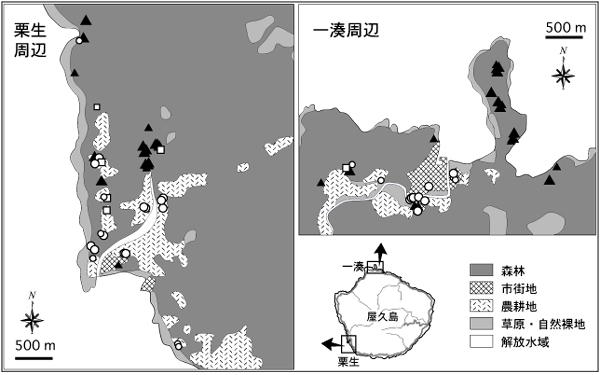
図2.栗生周辺と一湊周辺におけるヤモリ類の記録地点。シンボルの大小は、それぞれ確実な同定による記録と、暫定的な同定による記録を表す(本文参照)。▲ = ヤクヤモリ; ○ = ミナミヤモリ; □ = 交雑個体。土地利用区分のデータは環境省自然環境局生物多様性センターの生物多様性情報システムから入手し、幾つかの類似したカテゴリーを統合して図示した。

図3.ヤモリが記録された地点の土地利用タイプの比較。
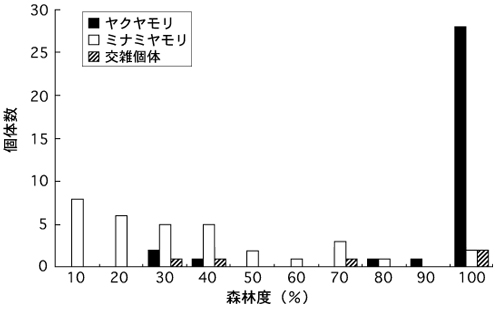
図4.ヤモリが記録された地点の周囲の森林度の比較。森林度は、各記録地点の周囲、半径100 mの区域に占める森林面積の割合
ミナミヤモリが主に集落や公園などの人為的に攪乱された環境に出現することは、2005年度と2006年度の調査で既に示されていた。ただし、そもそも屋久島のミナミヤモリが港を起点として集落に侵入・定着したのだとすると、屋久島全域を見たときにミナミヤモリの記録地点が集落やその近辺に存在する環境に偏るという結果はなかば自明であり、それによってこの種の分布を環境と直接関連づけることには問題があった。これに対し、今回の調査結果は、ミナミヤモリが定着している地域のなかでさえも2種のヤモリの出現環境が明確に異なることを示しており(図2,3)、これらのヤモリの分布がある程度、環境によって規定されていることが示唆された。すなわち、ミナミヤモリは屋久島に侵入後、集落や農耕地などに沿って選択的に分布を拡大させていると解釈することができる。
このことは、特に、栗生集落の北方の地域における2種の分布をみると分かりやすい。この地域では森林と農耕地が不連続に存在しているが、ヤモリの記録地点をプロットした地図からは、ヤクヤモリとミナミヤモリの出現が概ね環境と対応している様子が読み取れる(図2)。この結果は、ミナミヤモリが単に栗生集落を中心に分布を広げたわけではなく、人為的な攪乱を受けた環境に沿って分布拡大を果たしてきたことを強く示唆している。おそらく、市街地や農耕地といった環境はミナミヤモリの生息に適した何らかの条件を備えているのに対し、森林環境はむしろヤクヤモリの生息に適しているため、このような状況が生じるものと予想される。
とはいえ、栗生の北方において、ヤクヤモリが優占するはずの森林環境を越えて、隔離された農耕地パッチにもミナミヤモリが侵入していることを見逃すことはできない。一般に、ヤモリ類は物資の運搬に伴って人為的に持ち運ばれやすいため、これらの農耕地パッチのミナミヤモリが人手によって集落から直接運ばれたと考えることも可能かも知れない。しかし、栗生集落の北方にはいっさい集落がなく、人の往来も極めて限られている状況を考えると、この地域のミナミヤモリが人為的な持ち運びに由来するとは考えにくい。むしろ、少数個体が森林環境に侵入し、やがてはそれを越えて農耕地パッチに到達したと捉える方が自然であろう。この考えは、森林環境にも少数ながらミナミヤモリが出現するというバッファ解析の結果(図4)とも矛盾しない。
ここで、栗生を含む屋久島南西部の土地利用図を見ると、少なくとも栗生集落の南側では、集落の北方と同様に、農耕地が断続的に続いていることが分かる(図5a)。仮にミナミヤモリが森林環境を越えて分布拡大を果たせるとするならば、栗生のミナミヤモリは南側へも分布を拡大すると予想される。実際に過去の調査では、栗生のすぐ南側の中間集落から尾の大型鱗の状態がミナミヤモリの特徴と一致する個体が採集されており(柴田,1981)、さらにその南にある黒崎においてもミナミヤモリや,ミナミヤモリとヤクヤモリの交雑個体が発見されている(図1)。中間や黒崎は、島外との交易のある港が存在する地域から外れていることから、これらの地域のミナミヤモリの由来については説明が難しかったが、ミナミヤモリが狭い森林帯を越えて分散できると考えれば、その分布も無理なく説明できる。今後、この仮説を検証するため、中間や黒崎近辺の農耕地パッチにおけるヤモリ類の分布を詳しく調査する必要がある。
もう一つの調査地である一湊周辺においても、やはりミナミヤモリの記録はほぼ市街地と農耕地に限定されており、ヤクヤモリの出現環境との違いは明らかであった(図2)。このことは、ミナミヤモリが人為的攪乱を受けた環境に分布するという上述の議論と整合的である。しかし、一湊周辺では十分に密なデータが取れなかったことに加え、そもそも市街地・農耕地がほぼひと続きになっているため、ミナミヤモリが特定の環境に沿って分布拡大を果たしているかどうかについて厳密に議論することは難しい。
とはいえ、一湊周辺の森林地帯でもっぱらヤクヤモリばかりが記録された点は注目に値する(図2)。2005年度と2006年度の調査結果をみると、一湊のすぐ東側の森林地帯で1例だけミナミヤモリが記録されているものの、それを除けば一湊集落の外側で発見されたヤモリは全てヤクヤモリであり、さらに一湊と隣り合った志戸子や吉田の集落でさえもミナミヤモリは見つかっていない(図1)。これらの結果は、ミナミヤモリが、一湊から志戸子や吉田へは分散できていないことを示している。土地利用図をみると、一湊集落の周囲には隔離された農耕地パッチが存在せず、広い森林地帯によって最寄りの志戸子や吉田の集落と隔離されていることがわかる(図5b)。この状況は栗生地区とは対照的である。これらのことを総合すると、ミナミヤモリは、多少の森林地帯ならばそれを越えて分散するけれども、ヤクヤモリが優占する森林地帯の幅がある程度以上広くなると、もはやそれを越えて次の集落や農耕地パッチにたどり着くことはできないものと推測される。ただし、一湊と栗生の間にみられるミナミヤモリの分布様式の違いは、単に本種の侵入時期の違いを反映している可能性もあるため、この考えの妥当性については、今後の継続的な調査の結果に基づいて再度検討する必要がある。
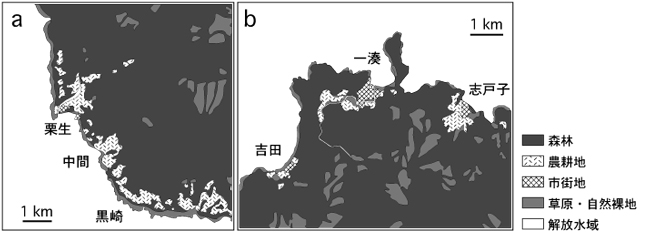
図5.(a)栗生を含む屋久島南西部の土地利用区分を示した地図と、(b)一湊を含む屋久島北部の土地利用区分を示した地図。土地利用区分のデータとカテゴリーは図2と同じ
今回の調査で確実に同定された栗生周辺の44個体と一湊周辺の34個体のうち、それぞれ4個体と1個体が交雑個体と判定された(表2)。つまり、栗生地区では、実に11個体に1個体が交雑個体と判定されたことになる。ここで、前述したように、今回の交雑個体の判定がその出現に関して過小評価する傾向にあることを考えれば、交雑個体の実際の出現率はさらに高い可能性もある。このように、少なくとも栗生近辺においては2種の交雑はかなり頻繁に起こっていると考えることができる。
交雑個体が記録された地点をみると、それは概ね、森林と市街地・農耕地の境界付近にあり(図2)、想定されるヤクヤモリとミナミヤモリの分布境界とほぼ一致する。この結果は、今回の基準で交雑個体と判定されるものが主に雑種第一代目の個体と予想されることを考えると理解しやすい。また、交雑個体の記録地点の環境をみると、2つの解析結果ともに、両親種のそれの中間的な傾向を示している(図3、4)。これも単純に、2種が出会いやすい場所で交雑が起こっていることを反映した結果とみなすことができる。
ここで、栗生の交雑個体4個体が、地理的にばらばらの地点で記録されている点にも注目したい(図2)。もし仮に、交雑がある特殊な条件下でのみ起こるのであれば、交雑個体は、その条件を満たす特定の地域で集中して発見されると期待されるが、実際の結果はそうなってはいない。さらに、過去の調査で発見された交雑個体の記録地点も各所に散在している(図1)。これらの状況から、ミナミヤモリとヤクヤモリの交雑はある特殊な条件下で起こるというよりも、むしろ2種が出会う機会があれば、至るところで起こるものと予想される。
前節の一湊地区に関する議論では、ミナミヤモリが広範囲な森林地帯には侵入できないとの予測を示した。しかし、交雑個体を含めて考えると、この見方には注意が必要である。上述したように、ミナミヤモリとヤクヤモリは累代で交雑し得るため(Toda et al., 2001)、そのような交雑を通してミナミヤモリ由来の遺伝子がヤクヤモリ集団のなかに深く浸透し、やがては森林地帯にまで到達する可能性がある。このような可能性を検討するため、今後、分子形質の分析を行って、戻し交雑を含む2種の交雑の進行状況を詳しく調査する必要がある。
ミナミヤモリが定着している栗生周辺と一湊周辺においてヤモリ類の分布を詳細に調べ、さらに地理情報システムを用いて、ヤモリの分布と環境との関係について検討した。その結果、ミナミヤモリは市街地や農耕地などの人為的攪乱を強く受けた環境に出現する傾向があり、ヤクヤモリは周囲を森林に囲まれた自然度の高い環境に出現することが分かった。特に、栗生の北方の地域では、森林によって隔てられた農耕地パッチでもミナミヤモリが確認されたことから、この種は集落や農耕地に沿って選択的に分布を広げてきたことが示唆された。この結果は、同時に、ミナミヤモリが狭い森林地帯を越えて分散できることも示している。また、これらの結果と地理情報に基づき、ミナミヤモリが栗生集落の南方へも分布を広げている可能性があることを示した。一方、周囲を広く森林に囲まれた一湊地区においては、ミナミヤモリの出現は集落近辺に限られていた。このことから、ミナミヤモリは、ヤクヤモリが優占する森林地帯が広く続いている状況では、それを越えて分散できないと考えられた。交雑個体は、特に栗生周辺で高率で発見され、2種の交雑が頻繁に起こっていることが示された。その記録地点は、想定される2種の分布境界と概ね一致し、出現環境も両親種のそれの中間的なものであった。これらの結果に加え、交雑個体が地理的に離れた幾つもの地点で発見されたことを考えると、2種の交雑はある特殊な条件下で起こっているというよりも、むしろ2種が出会いさえすれば比較的容易に起こると推測された。実際には、戻し交雑などを通して2種の間には深く遺伝子浸透が起こっている可能性もあるため、今回採取した組織片を用いて分子形質を分析し、2種の交雑についてさらに詳しく調べる必要がある。
一湊の小中学校の先生方には、校舎の周囲で調査することを許可していただいた。また、調査が夜間中心であったため、調査期間を通して屋久島フィールドワーク講座の他の班の講師、受講生の諸氏には生活面を中心にいろいろと便宜を図っていただいた。これらの方々に感謝の意を表します。
岡田純. 1998. 柴田保彦氏が採集した大隅半島周辺のヤモリ標本の検討. 比婆科学 (185): 7-13.
柴田保彦. 1981. 種子島のヤクヤモリ. 自然史研究1(15): 149-154.
竹内寛彦・戸田守. 2006. 馬毛島からのヤクヤモリの記録. 日本爬虫両生類学会報 2006: 26-27.
Toda, M., S. Okada, H. Ota, and T. Hikida. 2001. Biochemical assessment of evolution and taxonomy of the morphologically poorly diverged geckos, Gekko yakuensis and G. hokouensis (Reptilia: Squamata) in Japan, with special reference to their occasional hybridization. Biological Journal of the Linnean Society 73: 153-165.
京都大学野生動物研究センター > 屋久島フィールドワーク講座 > 第10回・2008年の活動−ヤモリ班−報告書
このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹