| 嫗搒戝妛栰惗摦暔尋媶僙儞僞乕亜壆媣搰僼傿乕儖僪儚乕僋島嵗亜戞9夞丒2007擭偺妶摦亅怉暔斍亅曬崘彂 |
| 奣梫 | 恖偲帺慠斍 | 僒儖斍 | 僔僇斍 | 怉暔斍 | 岞奐島嵗 | 僗僞僢僼 |
| 曬崘彂 | 姶憐暥 |
島巘丗懞忋揘柧丒悾旜柧峅
嶲壛幰丗堜忋懢庽丒戝媣曐枹嵷丒孠尨隳懢楴丒拞懞 桰
丂崱夞丄怉暔斍偱偼壆媣搰偺屌桳怉暔庬偺挷嵏傪峴偭偨丅庬丄垷庬丄曄庬傪崌寁偡傞偲栺1500偺暘椶孮偺僔僟怉暔偲庬巕怉暔偑壆媣搰偵惗堢偟偰偄傞偑丄偦偺偆偪壆媣搰屌桳偺怉暔庬偑47庬丄壆媣搰屌桳偺垷庬丒曄庬偑31暘椶孮偁傞偲偝傟偰偄傞乮搾杮丄2007乯丅偙傟傜偺壆媣搰屌桳怉暔偺拞偵偼丄嬤墢側怉暔庬偵斾傋偰怉暔懱偺僒僀僘偑彫宆壔偟偰偄傞傕偺偑彮側偔側偄丅偝傜偵丄偙偺傛偆偵彫宆壔偟偰偄傞屌桳怉暔偺懡偔偼壆媣搰偺崅抧偁傞偄偼宬棳偺嬤偔偵惗堢偟偰偄傞丅
丂偟偐偟丄崅抧傗宬棳増偄偺娐嫬偵偍偄偰壆媣搰偺彫宆壔偟偰偄傞條乆側屌桳怉暔偑屄懱儗儀儖偱偳偺掱搙彫宆壔偟偰偄傞偺偐丄偳偺傛偆偵偟偰怉暔懱偑彫偝偔側偭偰偄傞偺偐乮怉暔屄懱傪峔惉偟偰偄傞屄乆偺嵶朎偑彫宆壔偟偰偄傞偺偐丆嵶朎偺悢偑尭偭偰彫宆壔偟偰偄傞偺偐丆偦傟偲傕偦偺椉曽偑婲偙偭偰偄傞偺偐乯偼椙偔傢偐偭偰偄側偄丅摉慠偺偙偲側偑傜丄壆媣搰偺條乆側屌桳怉暔偱尒傜傟傞怉暔懱偺彫宆壔偑丆暋悢偺暘椶孮娫偱嫟捠偟偨婡峔偱偍偙偭偰偄傞偺偐丄傑偨丄暘椶孮偛偲偵堎側傞婡峔偱偍偙偭偰偄傞偺偐丄偲偄偆偙偲傕傢偐偭偰偄側偄丅偦偙偱丄崱夞偺挷嵏丒尋媶偱偼丄偄偔偮偐偺壆媣搰屌桳怉暔庬偵偮偄偰丄偦偺彫宆壔偲偄偆尰徾偑偳偺傛偆側婡峔偱偍偙偭偰偄傞偐丄偝傜偵偦偺傛偆側婡峔偼暘椶孮傪墇偊偰嫟捠側偺偐丆偦傟偲傕暘椶孮偛偲偵懡條側婡峔偱彫宆壔偟偰偄傞偺偐傪柧傜偐偲偡傞偙偲傪栚揑偲偟偨丅傑偨丄彫宆壔偟偰偄傞怉暔偑偳偺傛偆側娐嫬偵惗堢偟偰偄傞偐傕徻偟偔挷嵏偟偨丅
丂崱夞丄崅抧偵偍偄偰彫宆壔傪偟偰偄傞壆媣搰屌桳怉暔偲偟偰偼僀僢僗儞僉儞僇乮Solidago minutissima (Makino) Kitamura丂僉僋壢乯偲儎僋僔儅僯僈僫乮Ixeris parva (Kitamura) Yahara丂僉僋壢乯傪挷嵏丒尋媶偺懳徾偲偟偨乮恾1乯丅偙傟傜偺彫宆壔偟偨庬偲偺斾妑偵梡偄偨彫宆壔偟偰偄側偄嬤墢庬偼丄偦傟偧傟傾僆儎僊僜僂偲僯僈僫偱偁傞丅偨偩偟丆傾僆儎僊僜僂偼壆媣搰偱偼旕忢偵婬偱尒偮偗傞偙偲偑崲擄偱偁傝丄僯僈僫偼壆媣搰偵偼暘晍偟偰偄側偄丅偦偙偱庱搒戝妛搶嫗丒杚栰昗杮娰偵強憼偝傟偰偄傞僯僈僫偲傾僆儎僊僜僂偺墴偟梩昗杮傪梡偄偰屻弎偡傞宍懺宍幙偺寁應傪峴側偭偨丅偝傜偵丄僯僈僫偼嫗搒巗嵍嫗嬫偱嵦廤偟偨惗姅10屄懱傕斾妑娤嶡偵梡偄偨丅
丂堦曽丄掅抧偺宬棳増偄偺惗堢娐嫬偵偍偄偰彫宆壔偟偰偄傞怉暔偲偟偰僸儊僞僇僲僴僂儔儃僔乮Crypsinus yakushimensis (Makino) Tagawa丂僂儔儃僔壢乯側傜傃偵僒僀僑僋儂儞僌僂僔僟乮Lindsaea odorata Roxb. var. japonica (Bak.) Kramer丂儂儞僌僂僔僟壢乯傪挷嵏丒尋媶偺懳徾偲偟偨乮恾1乯丅偙傟傜俀庬偺僔僟怉暔偼壆媣搰屌桳偲偄偆傢偗偱偼側偄偑丄嬤墢庬偲峫偊傜傟偰偄傞僞僇僲僴僂儔儃僔Crypsinus engleri (Luerss.) Copel.偍傛傃儂儞僌僂僔僟L. odorata Roxb.偲斾傋傞偲怉暔懱偼旕忢偵彫宆壔偟偰偄傞乮娾捨 1992乯丅偝傜偵丄壆媣搰偺宬棳増偄偺娐嫬偱偼挊偟偔彫宆壔偟偨屄懱傕懡悢尒傜傟傞庬偱偁傞丅
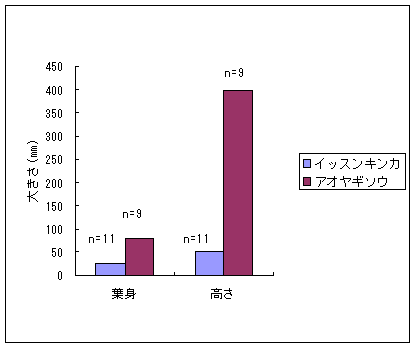
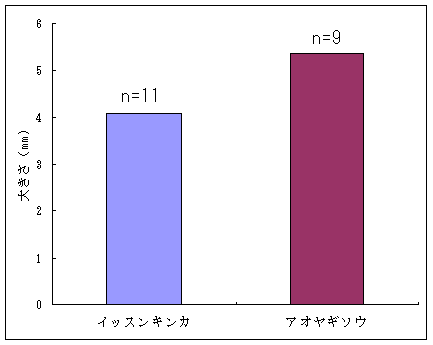
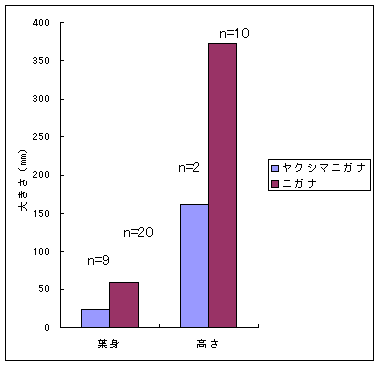
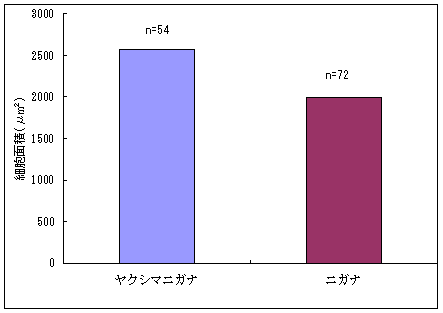
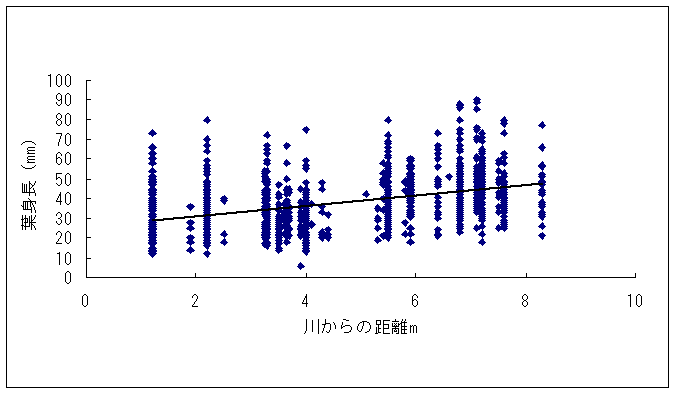
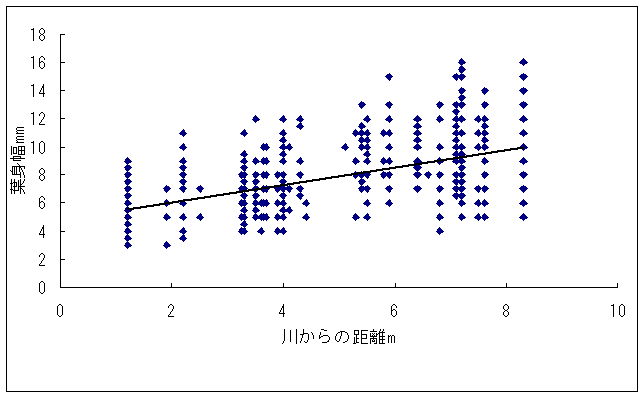
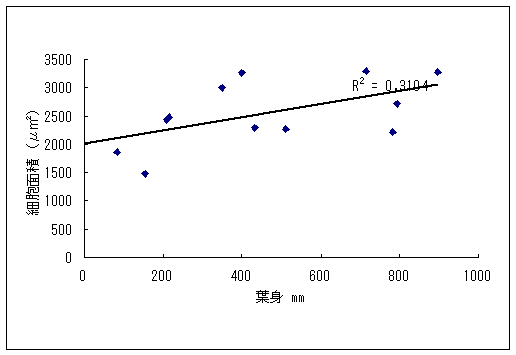
恾1丂僀僢僗儞僉儞僇乮忋嵍乯丄儎僋僔儅僯僈僫乮忋拞乯丄僸儊僞僇僲僴僂儔儃僔乮忋塃乯丄僒僀僑僋儂儞僌僂僔僟乮壓乯 僀僢僗儞僉儞僇偍傛傃儎僋僔儅僯僈僫偼壆媣搰偺崅抧偵偟偐惗堢偟偰偄側偄偺偱丄8寧22擔偵崟枴妜偐傜壴擵峕壨廃曈乮昗崅1600-1830m乯偱嵦廤傪偟偨丅偦傟偧傟偺怉暔庬偵偮偄偰16屄懱丄10屄懱偺僒儞僾儖傪嵦廤偟丄屄懱偛偲偵堎側傞斣崋傪偮偗偨價僯乕儖戃偵擖傟偰帩偪婣偭偨丅崱夞嵦廤偟偨僀僢僗儞僉儞僇偼慡偰奐壴偟偰偄傞屄懱偱偁傞丅偟偐偟丄儎僋僔儅僯僈僫偵偮偄偰偼丄奐壴偟偰偄傞屄懱偼1屄懱偟偐嵦廤偡傞偙偲偼偱偒側偐偭偨丅帩偪婣偭偨僒儞僾儖偼應掕偵梡偄傞傑偱丄椻憼屔乮係亷乯偱曐娗偟偨丅 丂愭偵傕弎傋偨傛偆偵僸儊僞僇僲僴僂儔儃僔偲僒僀僑僋儂儞僌僂僔僟偼丄壆媣搰偺宬棳増偄偱挊偟偔彫宆壔偟偨屄懱偑懡悢尒傜傟傞僔僟怉暔偱偁傞丅偦偙偱丄巹偨偪偼壆媣搰偺壴梘愳増偄乮昗崅 栺180m乯偵惗堢偟偰偄傞僸儊僞僇僲僴僂儔儃僔偍傛傃僒僀僑僋儂儞僌僂僔僟偺廤抍偵偍偄偰丄偳偺傛偆側婡峔偱怉暔懱偑彫宆壔偟偰偄傞偐丄偦偟偰惗堢娐嫬偲怉暔懱僒僀僘偺娭學偑偳偺傛偆偵側偭偰偄傞偐傪柧傜偐偵偡傞偙偲傪栚揑偲偟偨挷嵏傪峴偭偨丅 恾2丂儔僀儞丒僩儔儞僛僋僩朄偱婰榐偟偨壴梘愳偵偍偗傞僸儊僞僇僲僴僂儔儃僔偺屄懱孮偺埵抲 丂怉暔懱偺抧忋晹偺崅偝偲梩偺戝偒偝偺暯嬒抣偼丄僀僢僗儞僉儞僇偱偼偦傟偧傟5.00cm偲27.55mm偱偁偭偨偺偵懳偟偰丄傾僆儎僊僜僂偱偼偦傟偧傟39.80cm偲79.67mm偱偁偭偨 乮恾3乯丅僀僢僗儞僉儞僇偼傾僆儎僊僜僂偵斾傋偰怉暔懱偑旕忢偵彫偝偔側偭偰偄偨丅偲偙傠偑壴偺戝偒偝偺暯嬒抣偼丄僀僢僗儞僉儞僇偺4.07mm偵懳偟偰丄傾僆儎僊僜僂偼5.36mm偲側傝丄傗傗僀僢僗儞僉儞僇偺壴偺傎偆偑彫偝偐偭偨傕偺偺丄怉暔懱偺崅偝偱尒傜傟偨傛偆側戝偒側堘偄偼側偐偭偨 乮恾 4乯丅偝傜偵丄梩偺嵶朎偺戝偒偝偺暯嬒抣偼丄僀僢僗儞僉儞僇偺1061.09?m2偵懳偟偰丄傾僆儎僊僜僂偱偼1035.40?m2偲側傝 乮恾 5丄6乯丄嵶朎偺戝偒偝偱偼僀僢僗儞僉儞僇偲傾僆儎僊僜僂偺娫偱傎偲傫偳嵎偼尒傜傟側偐偭偨丅
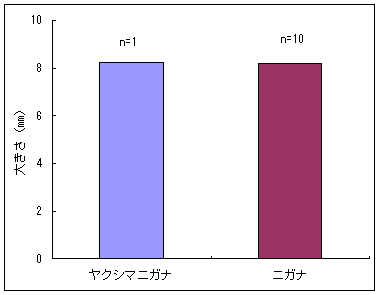
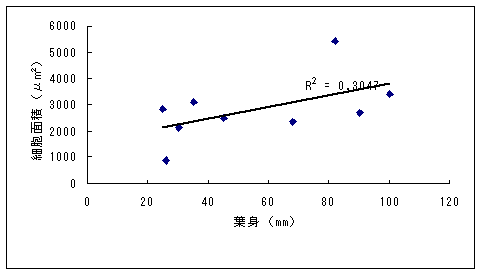
恾3丂僀僢僗儞僉儞僇偲傾僆儎僊僜僂偺梩恎偺挿偝偲怉暔懱偺崅偝 恾4丂僀僢僗儞僉儞僇偲傾僆儎僊僜僂偺壴偺戝偒偝乮憤錃曅偺挿偝乯 恾5-1丂僀僢僗儞僉儞僇乮屄懱no. 3乯偺梩偺昞旂嵶朎 恾5-2丂傾僆儎僊僜僂乮昗杮斣崋5325乯偺梩偺昞旂嵶朎偲偦偺戝偒偝 恾6丂僀僢僗儞僉儞僇乮16屄懱乯偲傾僆儎僊僜僂乮4屄懱乯偺梩昞旂嵶朎偺柺愊偺暯嬒抣 丂怉暔懱偺崅偝偍傛傃梩偺戝偒偝偺暯嬒抣偼丄儎僋僔儅僯僈僫偱偼偦傟偧傟16.05cm丄22.89mm偱偁偭偨偺偵懳偟偰丄僯僈僫偱偼偦傟偧傟37.26cm丄59.25mm偱偁偭偨乮恾 7乯丅儎僋僔儅僯僈僫偼僯僈僫偵斾傋偰怉暔懱偑偼傞偐偵彫偝偔側偭偰偄偨丅堦曽丄壴偺戝偒偝偺暯嬒抣偼丄儎僋僔儅僯僈僫偱偼8.21mm偵懳偟偰丄僯僈僫偱偼8.19mm偱偁傝乮恾 8乯丄傎偲傫偳堘偄偼尒傜傟側偐偭偨丅傑偨丄梩偺嵶朎偺戝偒偝偺暯嬒抣偼丄儎僋僔儅僯僈僫偱偼2570.80?m2偵懳偟偰丄僯僈僫偱偼1995.94?m2偱偁傝丄儎僋僔儅僯僈僫偺嵶朎僒僀僘偺傎偆偑戝偒偐偭偨乮恾 9丄10 乯丅偮傑傝丄怉暔懱儗儀儖偱偼傛傝彫偝偐偭偨儎僋僔儅僯僈僫偺傎偆偑丄媡偵梩偺嵶朎偼傛傝戝偒偐偭偨丅
恾7丂儎僋僔儅僯僈僫偲僯僈僫偺梩恎偺挿偝偲怉暔懱偺崅偝
恾8丂儎僋僔儅僯僈僫偲僯僈僫偺壴偺戝偒偝乮憤錃曅偺挿偝乯 恾9-1丂儎僋僔儅僯僈僫乮屄懱斣崋1乯偺梩偺昞旂嵶朎偲偦偺戝偒偝 恾9-2丂僯僈僫乮屄懱斣崋7乯偺梩偺昞旂嵶朎偺戝偒偝 恾10丂儎僋僔儅僯僈僫乮6屄懱乯偲僯僈僫乮9屄懱乯偺梩昞旂嵶朎偺柺愊偺暯嬒抣 丂壴梘愳廤抍偱朎巕傪偮偗偰惉弉偟偰偄傞偲峫偊傜傟傞1069屄懱偺梩偺戝偒偝乮梩恎挿偲梩恎暆乯傪應掕偟偨丅愳偐傜偺嫍棧偑棧傟傞偵偮傟偰梩恎挿偼挿偔丄梩恎暆偼嵶偔側偭偰偄偭偨乮恾 11丄12乯丅傑偨丄愳偺拞偵偁傞娾忋偺屄懱偺梩偼旕忢偵嵶偐偭偨丅堦曽丄梩偺嵶朎偺戝偒偝傕丄梩偺僒僀僘乮梩恎挿亊梩恎暆乯偺彫偝側屄懱偱偼丄梩僒僀僘偺戝偒側屄懱偵斾傋偰彫偝偔側偭偰偄偨乮恾 13乯丅偡側傢偪丄僸儊僞僇僲僴僂儔儃僔偼嵶朎僒僀僘偑彫偝偔側傞偙偲偱怉暔懱偑彫宆壔偟偰偄偨丅
丂僸儊僞僇僲僴僂儔儃僔偲摨條偵梩偺挿偝偑抁偄屄懱傎偳丄梩偺嵶朎偺僒僀僘傕彫偝偐偭偨乮恾14乯丅僒僀僑僋儂儞僌僂僔僟偼丄嵶朎傪彫偝偔偡傞偙偲偵傛偭偰怉暔懱傪彫宆壔偝偣偰偄傞偙偲偑柧傜偐偵側偭偨丅傑偨丄傛傝彫宆偺梩傪傕偮屄懱傎偳丄傛傝愳偺棳傟偵嬤偄応強偵惗堢偟偰偄偨丅
丂崅抧偵惗堢偟偰偄傞旐巕怉暔偱偁傞僀僢僗儞僉儞僇偍傛傃儎僋僔儅僯僈僫偺梩偺僒僀僘側傜傃偵怉暔懱偺崅偝偼丄掅抧偵惗堢偟偰偄傞傾僆儎僊僜僂偍傛傃僯僈僫偺怉暔懱僒僀僘偵斾傋偰彫宆壔偟偰偄偨丅堦曽丄惗怋婍姱偱偁傞壴偺僒僀僘偵偮偄偰偼丄偙傟傜崅抧偲掅抧偵偦傟偧傟惗堢偡傞嬤墢側怉暔庬偺娫偱丄塰梴婍姱偱尒傜傟偨傎偳偺堘偄偼側偐偭偨丅傑偨塰梴婍姱偺梩傪峔惉偟偰偄傞嵶朎偺戝偒偝偵偮偄偰傕丄偙傟傜崅抧偲掅抧偺嬤墢庬娫偱嵎偑側偄偐丄偁傞偄偼儎僋僔儅僯僈僫偺傛偆偵媡偵崅抧偱彫宆壔偟偰偄傞庬偺曽偑梩偺嵶朎偼傛傝戝偒偔側偭偰偄偨丅偄偢傟偵偟偰傕梩偺嵶朎偼彫偝偔偼側偭偰偼偄側偐偭偨偺偱丄僀僢僗儞僉儞僇偲儎僋僔儅僯僈僫偱偼丄梩側偳偺塰梴婍姱偺嵶朎悢傪尭彮偝偣傞偙偲偵傛偭偰怉暔懱傪彫宆壔偝偣偨偲峫偊傜傟傞丅傑偨丄塰梴婍姱偵偮偄偰偼丄僀僢僗儞僉儞僇偲儎僋僔儅僯僈僫偼嫟捠偟偨婡峔偱彫宆壔偝偣偰偄偨偲傕偄偊傞丅 丂壆媣搰偺崅抧偵偍偄偰彫宆壔偟偰偄傞旐巕怉暔偱偁傞僀僢僗儞僉儞僇偍傛傃儎僋僔儅僯僈僫偼丄偲傕偵塰梴婍姱偱偁傞梩偺嵶朎悢傪尭傜偟偰彫宆壔偟偰偄傞偙偲偑柧傜偐偲側偭偨丅旐巕怉暔偺梩偺敪惗偺夁掱偵偍偄偰偼丄嵶朎暘楐偑妶敪偵婲偙偭偰嵶朎悢偑憹偊傞嵶朎憹怋婜偲丄嵶朎暘楐偼婲偙傜偢偵屄乆偺嵶朎偑怢挿偡傞嵶朎奼戝婜偺2偮偺帪婜偑偁傞乮壛摗丄1999乯丅僀僢僗儞僉儞僇側傜傃偵儎僋僔儅僯僈僫偱偼梩偺嵶朎悢傪尭傜偟偰偍傝丄偙偺偙偲偼梩偺宍惉夁掱偵偍偄偰嵶朎暘楐偺夞悢傪尭傜偟偰偄傞偙偲偵側傞丅偟偨偑偭偰丄偙傟傜偺旐巕怉暔庬偼嵶朎憹怋婜傪抁弅偝偣偰偄傞壜擻惈偑峫偊傜傟傞乮奣嶼偱嵶朎悢偼掅抧偺傕偺偲斾傋偰1/20偵側偭偰偄傞乯丅 娾捨朚抝. 1992. 擔杮偺栰惗怉暔僔僟. 暯杴幮, 搶嫗. 嫗搒戝妛栰惗摦暔尋媶僙儞僞乕 亜 壆媣搰僼傿乕儖僪儚乕僋島嵗
亜戞9夞丒2007擭偺妶摦亅怉暔斍亅曬崘彂 偙偺儁乕僕偺栤偄崌傢偣愭丗嫗搒戝妛栰惗摦暔尋媶僙儞僞乕丂悪塝廏庽丂
僀僢僗儞僉儞僇偲儎僋僔儅僯僈僫
丂屄懱儗儀儖偺僒僀僘傪斾妑偡傞偨傔偵怉暔懱偺抧忋晹偺崅偝乮抧柺偐傜嵟忋晹偺壴傑偱偺崅偝乯丄梩乮奺屄懱偱嵟戝偺宻梩乯偺挿偝傪掕婯傪梡偄偰應掕偟偨丅壴偺僒僀僘乮憤錃曅偺挿偝乯偼僨僕僞儖尠旝嬀乮僉乕僄儞僗幮惢乯傪梡偄偰娤嶡偲寁應偺椉曽傪峴偭偨丅
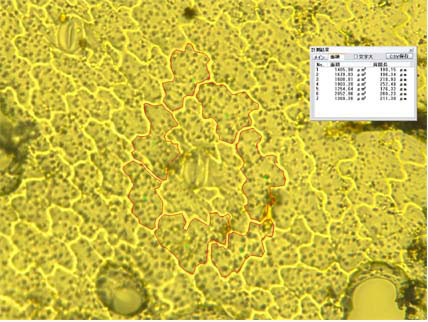
丂嵶朎儗儀儖偺僒僀僘傪斾妑偡傞偨傔偵梩偺昞旂嵶朎偺戝偒偝傪應掕偟偨丅應掕偵偼丄傑偢僗儞僾朄乮Suzuki乫s Universal Micro-Printing Method乯偵傛傝梩偺棤柺偺昞旂嵶朎孮偺儗僾儕僇傪偲偭偨丅偦傟傪僨僕僞儖尠旝嬀梡偄偰娤嶡偟丄婥岴傪偲傝傑偔嵶朎孮乮岴曅嵶朎偍傛傃岴曅嵶朎偵愙偟偰偄傞嵶朎傪彍偔乯傪峔惉偡傞嵶朎侾偮侾偮偺柺愊偺應掕傪偟丄偦傟傜偺暯嬒抣傪偦偺屄懱偺梩偺嵶朎僒僀僘偲偟偨丅
僸儊僞僇僲僴僂儔儃僔偲僒僀僑僋儂儞僌僂僔僟
丂挷嵏抧揰偵挷嵏帪偺悈棳偺抂偐傜悈偺棳傟偲悅捈曽岦偵悈棳懁偵2m丄娸懁偵8m丄崌寁偟偰10m儔僀儞偑偱偒傞傛偆偵50倣偺姫偒広傪愝抲偟丄儔僀儞偐傜忋棳懁丒壓棳懁偵偦傟偧傟3m偺斖埻偺儔僀儞丒僩儔儞僛僋僩傪愝掕偟偨 乮恾 2乯丅偦偟偰丄愝掕偟偨僩儔儞僛僋僩撪偺慡屄懱偺僸儊僞僇僲僴僂儔儃僔偺惉弉梩偺梩恎挿偲梩恎暆傪掕婯偱寁應偟偨丅僸儊僞僇僲僴僂儔儃僔偼娾偺忋偵惗堢偟偰偄傞偺偱丄惗偊偰偄偨娾偺埵抲傪僩儔儞僛僋僩忋偵婰榐偟偨丅偝傜偵丄梩偺戝偒偝偺堘偄偵傛偭偰梩偺嵶朎偺戝偒偝偑偳偺傛偆偵側偭偰偄傞偐寁應偡傞偨傔偵丄偙偺廤抍拞偱摿偵彫宆偺屄懱偍傛傃戝宆偺屄懱偐傜丄偦傟偧傟3屄懱偢偮傪嵦廤傪偟丄價僯乕儖戃偵擖傟偰帩偪婣偭偨丅僒僀僑僋儂儞僌僂僔僟偼儔僀儞丒僩儔儞僛僋僩撪偵惗堢偟偰偄傞條乆側戝偒偝偺怉暔懱傪丄偦傟偧傟偺戝偒偝偛偲偵暘偗偰價僯乕儖戃偵擖傟偰帩偪婣偭偨丅
丂帩偪婣偭偨僸儊僞僇僲僴僂儔儃僔偍傛傃僒僀僑僋儂儞僌僂僔僟偼丄僀僢僗儞僉儞僇傗儎僋僔儅僯僈僫偲摨條偺曽朄偱丄偦偺梩偺嵶朎偺戝偒偝傪寁應偟偨丅

寢壥
僀僢僗儞僉儞僇偲傾僆儎僊僜僂

(慄偱埻傑傟偨嵶朎偺柺愊傪僨僕僞儖尠旝嬀傪梡偄偰寁應偟偨)
(慄偱埻傑傟偨嵶朎偺柺愊傪僨僕僞儖尠旝嬀傪梡偄偰寁應偟偨)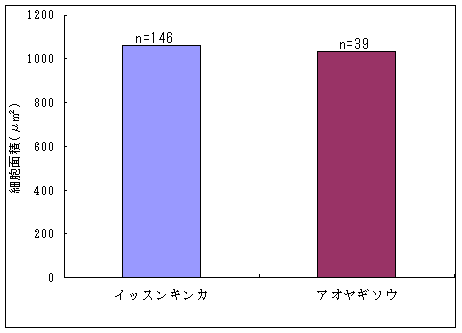
乮n偼寁應偟偨嵶朎悢偺崌寁乯儎僋僔儅僯僈僫偲僯僈僫

乮n偼寁應偟偨嵶朎悢偺崌寁乯僸儊僞僇僲僴僂儔儃僔
恾11丂僸儊僞僇僲僴僂儔儃僔偑惗堢偟偰偄偨応強偺愳偐傜偺嫍棧偲偦偺梩恎挿偲偺娭學
恾12丂僸儊僞僇僲僴僂儔儃僔偑惗堢偟偰偄偨応強偺愳偐傜偺嫍棧偲偦偺梩恎暆偲偺娭學
恾13丂僸儊僞僇僲僴僂儔儃僔偺梩偺戝偒偝乮梩恎挿x梩恎暆乯偲梩偺昞旂嵶朎偺戝偒偝偲偺娭學僒僀僑僋儂儞僌僂僔僟
恾14丂僒僀僑僋儂儞僌僂僔僟偺梩偺戝偒偝乮梩恎挿乯偲梩偺昞旂嵶朎偺戝偒偝偲偺娭學峫嶡
丂堦曽丄宬棳増偄偲偄偆娐嫬曄摦偺戝偒側応強偵惗堢偟偰偄傞僔僟怉暔偱偁傞僸儊僞僇僲僴僂儔儃僔偱偼丄愳偺棳傟偐傜棧傟傞偵偟偨偑偭偰丄梩偺僒僀僘偑戝偒偔側傞孹岦偑尒傜傟偨丅僸儊僞僇僲僴僂儔儃僔偼梩傪彫宆壔偝偣傞偙偲偵傛偭偰丄愳偑憹悈偟偨偲偒偵偝傜偝傟傞棳悈掞峈傪尭偠偰丄梩傪堷偒偪偓傜傟側偔偡傞揔墳傪偟偰偄傞傕偺偲峫偊傜傟傞丅偝傜偵僸儊僞僇僲僴僂儔儃僔偺梩偺彫偝側屄懱偼丄戝偒側屄懱傛傝傕梩偺嵶朎僒僀僘偑彫偝偔側偭偰偄偨丅偝傜偵丄宬棳増偄偺娐嫬偵僸儊僞僇僲僴僂儔儃僔偲偟偽偟偽崿惗偟偰惗堢偟偰偄傞僒僀僑僋儂儞僌僂僔僟偵偮偄偰傕丄梩偺傛傝彫偝側屄懱偼戝偒側屄懱偵斾傋偰丄梩偺嵶朎僒僀僘偑彫偝偔側偭偰偄偨丅偡側傢偪丄宬棳増偄偵惗堢偡傞僔僟怉暔偱偼梩偺嵶朎偑彫偝偔側傞偙偲偱梩傪彫宆壔偝偣偰偄傞偙偲偑嫟捠偟偰娤嶡偝傟偨丅偟偨偑偭偰丄宬棳増偄偺僔僟怉暔偵尒傜傟偨彫宆壔偺婡峔偼堎側傞暘椶孮娫偱嫟捠偟偰偄偨丅堦曽偱丄崅抧偱彫宆壔偟偰偄傞旐巕怉暔偲宬棳増偄偱彫宆壔偟偰偄傞僔僟怉暔偼丄堎側傞婡峔偵傛偭偰梩傪彫宆壔偝偣偰偄傞偙偲傕柧傜偐偵側偭偨丅
傑偲傔
丂偦偺堦曽偱丄惗怋婍姱偱偁傞壴偺僒僀僘偱偼丄崅抧偵惗堢偟偰怉暔懱偑傛傝彫宆壔偟偰偄傞僀僢僗儞僉儞僇偍傛傃儎僋僔儅僯僈僫偼丄掅抧偵惗堢偡傞傾僆儎僊僜僂側傜傃偵僯僈僫偲斾妑偟偰丄傎偲傫偳摨偠僒僀僘偱偁偭偨丅偙偺偙偲偼憲暡崺拵偺懱僒僀僘偼丄崅抧偱傕掅抧偱傕偦傟傎偳戝偒偔曄傢偭偰偍傜偢丄崅抧偵偍偄偰壴僒僀僘傪梩僒僀僘偲摨偠傛偆偵彫偝偔偟偰偟傑偆偲朘壴崺拵偵憲暡傪偟偰傕傜偊側偔側傝丄揔墳搙偑掅壓偟偰偟傑偆偨傔偱偼側偄偐偲峫偊傜傟偨丅
丂偝傜偵丄儎僋僔儅僯僈僫偱偼梩偺嵶朎悢傪尭傜偡偙偲偩偗偱側偔丄梩偺嵶朎傪傛傝戝宆壔偝偣偰偄傞偲偄偆嫽枴怺偄寢壥傕摼傜傟偨丅偡側傢偪儎僋僔儅僯僈僫偱偼丄嵶朎憹怋婜娫傪抁弅偝偣傞堦曽丄嵶朎奼戝婜偼僯僈僫傛傝傕挿偔側偭偨壜擻惈偑峫偊傜傟傞丅偙偺嫽枴怺偄尰徾偺恑壔妛揑丄揔墳揑堄媊偼傑偩椙偔傢偐傜側偄偑丄屄乆偺嵶朎僒僀僘偺戝宆壔偼儎僋僔儅僯僈僫偺怉暔懱偺嫮搙傪庛偔偡傞偙偲偵側傝偐偹側偄偑丄怉暔懱慡懱偑彫宆壔偡傞偙偲偱晽側偳偺塭嬁偑彫偝偔側傝丄怉暔懱偺嫮搙偑壓偑偭偰傕戝偒側栤戣偑側偄偺偐傕抦傟側偄丅崱屻丄儎僋僔儅僯僈僫偺梩偺敪惗夁掱偵偮偄偰偼丄傛傝徻嵶側尋媶偑朷傑傟傞丅
丂堦曽丄壆媣搰偺宬棳増偄偺娐嫬偵惗堢偡傞俀偮偺僔僟怉暔庬偱娤嶡偝傟偨梩偺彫宆壔偺婡峔偼丄嵶朎僒僀僘傪彫偝偔偡傞偲偄偆傕偺偱偁偭偨丅僔僟怉暔偺梩偺敪惗偵偍偄偰傕丄嵶朎暘楐婜偲怢挿婜偑偁傞偙偲偑抦傜傟偰偄傞丅偙傟傜偺宬棳増偄偺僔僟怉暔庬偱偼丄梩偺宍惉夁掱偵偍偄偰嵶朎憹怋婜偼曄壔偟偰偄側偄偑丄嵶朎怢挿婜偑抁弅偝傟偰偄傞壜擻惈偑峫偊傜傟傞丅搶撿傾僕傾嶻偺宬棳増偄僔僟怉暔庬偵偍偄偰丄嬤墢側棨忋庬偲斾妑偡傞偲昞旂嵶朎偺僒僀僘偑彫偝偔側偭偰偄傞偙偲偑曬崘偝傟偰偄傞乮壛摗丄1999乯丅偮傑傝丄嵶朎僒僀僘偑彫偝偔側偭偰梩偑彫宆壔偡傞丄偁傞偄偼梩偑嵶偔側傞偙偲偼丄宬棳増偄僔僟怉暔偱偼惗堢偡傞抧堟傪墇偊偰嫟捠偟偰傒傜傟傞偺偱偁傞丅
丂崱夞偺挷嵏偺寢壥丄崅抧偵偍偗傞旐巕怉暔偺彫宆壔偼嵶朎悢偺尭彮偵傛偭偰丄堦曽丄宬棳増偄偺僔僟怉暔偱偼嵶朎僒僀僘偺彫宆壔偵傛偭偰丄偦傟偧傟怉暔懱僒僀僘傪彫偝偔偝偣偰偄傞偙偲偑柧傜偐偲側偭偨丅偟偨偑偭偰丄摨偠彫宆壔偡傞偲偄偆尰徾偱偼偁偭偰傕丄暘椶孮偺堘偄丄偁傞偄偼惗堢偡傞娐嫬偺堘偄偵傛偭偰彫宆壔偡傞婡峔偑堎側偭偰偄偨偲偄偆傢偗偱偁傞丅堦尒摨偠傛偆偵尒偊傞尰徾偑丄慡偔堎側傞婡峔偱婲偙偭偰偄傞偙偲偑柧傜偐偵側偭偨偙偲偼旕忢偵嫽枴怺偄丅
堷梡暥專
壛摗夒孾. 1999. 怉暔偺恑壔宍懺妛. 搶嫗戝妛弌斉夛, 搶嫗.
搾杮婱榓. 2007. 怉暔偺曮屔丒壆媣搰. 廐摴抭淺乮曇乯, 悈偲悽奅堚嶻, pp. 93-107, 彫妛娰, 搶嫗.