| 京都大学野生動物研究センター>屋久島フィールドワーク講座>第8回・2006年の活動-サル班-報告書 |
| 概要 | 人と自然班 | サル班 | シカ班 | ヤモリ班 | 植物班 | 公開講座 | スタッフ |
| 報告書 | 感想文 |
講師: 中川尚史、松原幹
参加者: 小沢友理子、増山遥、住栄貴恵、野村知代、下中麻奈美
近年屋久島の半山では、ヤクシカとヤクシマザルが同時に観察されることが多い。そして西部林道ではシカの増加が原因のひとつと考えられている林床植生の貧困化に伴い、シカの採食が困難になっていると思われる。
このためシカがサルから特に採食に関して何らかの利益を得るようになり、シカとサルの関係が以前より緊密になったためシカとサルが同時に観察されるようになったのではないだろうか。
この考えのもと、我々はシカとサルの種間関係について調査を行った。
ヤクシマザルの半山E群(計29頭)
ヤクシカのダニー(オス2歳)
西部林道半山地区(南稜~北稜)
2006年8月23日~8月27日
1)サル観察(学生3名+指導者
・観察者A
E群の群れを追跡。極力多くの群れのメンバーが遊動する方向についていき、彼らが樹上採食を開始したら、樹木種、樹木個体名(番号)とともに、5分毎に採食頭数を採食終了時(サルが全ていなくなった時)まで記録した。
・観察者B
GPSを携行し、位置データを1分毎に自動記録するとともに、5分間隔でDOP値と未受信 / 2D / 3Dの別を記録した。その際、視界内のシカの有無、有の場合はシカと樹下のサルの頭数と採食の有無を、樹下と樹外に分けて記録した。サルとシカの最接近固体同士の距離を目測して記録した。
・観察者C
サルが採食樹に上がると同時に樹下を掃除した。全ては不可能なので1m四方の方形区を樹下に設置し、その中の古い落果/落葉は全て除去。ただし、今回のサルの訪問によって落ちた新しい落果/落葉は数えたのち方形区内に戻した。
この方形区内の落果/落葉重量をシカの採食開始時と採食終了時において計測し、シカの採食量を調べた。
またこの際、採食樹の樹種、樹種ナンバー、樹冠の直径と短径も測定記録した。
2)シカ観察(学生2名+指導者)
観察者D
特定のシカを追跡。採食を開始したら、採食品目とともに採食頭数を5分毎に記録。落葉/落果採食の場合には、採食頭数は樹下とそれ以外に分け、さらに樹木個体名も記録した。
観察者E
GPSを携行し、位置データを1分毎に自動記録するとともに、5分間隔でDOP値と未受信 / 2D / 3Dの別を記録した。その際、視界内のサルの有無、有の場合はそのサルの頭数と採食の有無を樹下、樹上、樹外に分けて記録した。サルとシカの最近接個体同士の距離を目測して記録した。
指導者
サル観察の観察者Cと同様。
シカ観察班の観察者Dがシカ群から少し離れたところに移動する。そこでスピーカーを設置し、以下の条件でサルの集合採食時のクーコールを再生した。
1)音声のみ(地上)、2)音声のみ(樹上)、3)葉音のみ、4)葉音+音声
実験対象は1)短角(1歳♂)2)ダニー(2歳♂)3)ダニエル(2歳♂)4)6チャン(3歳♂)
観察者Eはシカの反応、1)首を上げ(耳を動かす)、2)音源を注視する、3)音源の方に移動するなどを記録した。
サルが道路上でした糞の位置をプロットし、チョークで囲って番号をつけて識別しておいた。観察中にシカにより採食されれば、確認時刻とともに記録し、サルが去った後も消失をチェックできるようにした。少なくとも、その日の実験終了時には糞の有無を確認時刻とともに再チェックした。その際、糞の消失あるいは分解の程度を記録し、それがシカによるものなのか、あるいはそれ以外(フンコロガシなど)によるものなのかを記録した。
(下中 麻奈美)
シカが直接採取出来ない高さの木から葉や果実をシカが採食することが頻繁に観察された。しかし、樹種によってシカが好むもの、好まないものとが大きく分かれる。
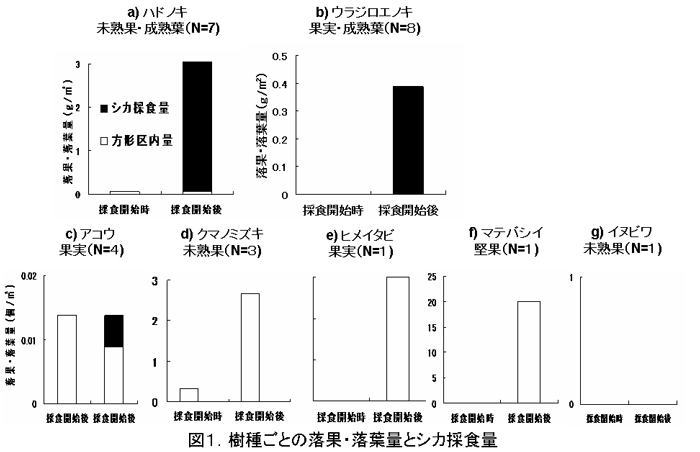
ハドノキおよびウラジロエノキはサルによる落下果実、葉のほとんどを、アコウは落下果実量の3分の1程度がシカによって採食されており、クマノミズキ、ヒメイタビ、マテバシイにおいては落下果実はまったく採食されなかった。イヌビワに関しては、サルの採食時に果実が地上に落とされることはなかったということが示されている。(図1)
図1.樹種ごとの落果・落葉量とシカ採食量
図2は、サルが樹上にいるとき樹下にいるシカのうちどのくらいの割合のものが採食行動をしているかを示している。3日間の調査により、1日目は80%、2日目は85.7%、3日目は91.7%、平均85.8%のシカが樹下で採食行動をしているという結果が得られた。 このようにシカが樹下において高い割合で採食行動をしている理由にはシカがサルの採食に際する落下物を求めて集まってきていることが考えられ、サルはシカの採食行動に大きな影響を与えていると結論づけることができる。
(小澤 友理子)
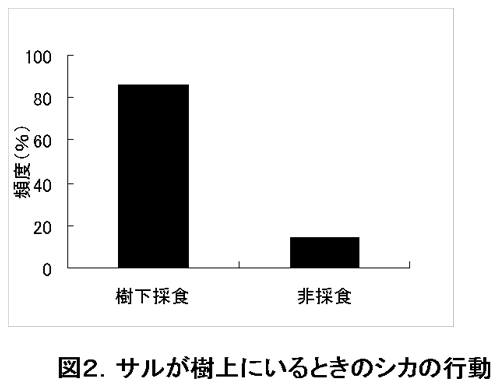
図2.サルが樹上にいるときのシカの行動
サルを利用するために、シカは採食場所をどのように選択しているのだろうか。
ここで、ある特定の採食樹における、サルとシカの採食頭数の時間推移による変化を調べた(図3~5)。
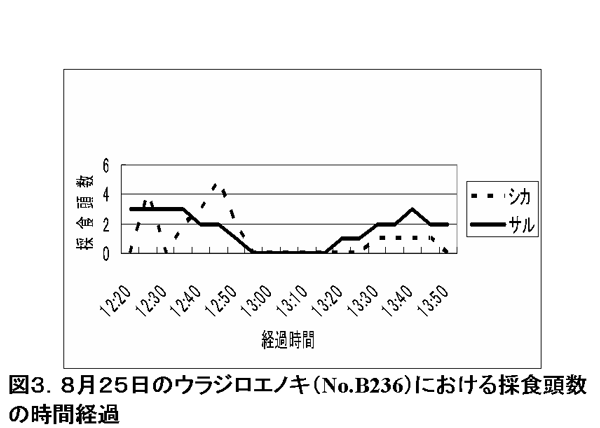
図3. 8月25日のウラジロエノキ(No.B236)における採食頭数の時間経
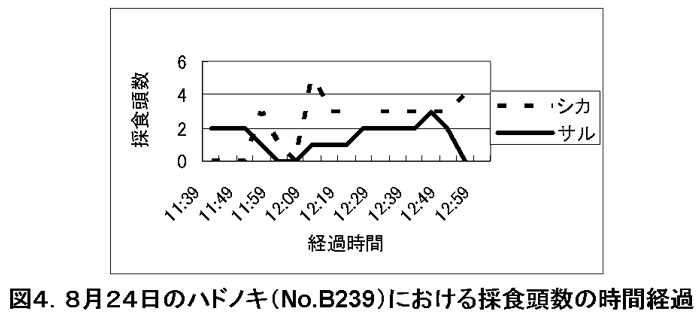
図4. 8月24日のハドノキ(No.B239)における採食頭数の時間経
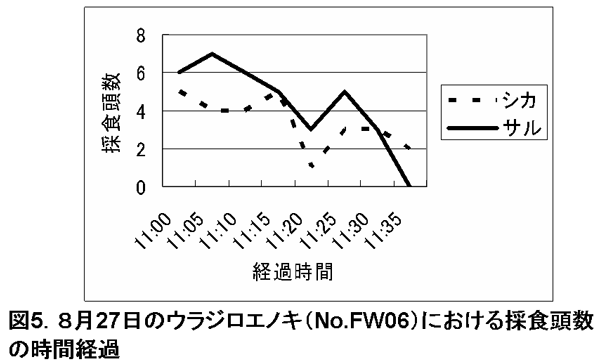
図5. 8月27日のウラジロエノキ(No.FW06)における採食頭数の時間経
調査期間中、特に採食の利用が多かったのは、ウラジロエノキとハドノキであった。これらの樹種では、サルが樹上採食を始めた後、その樹下にシカが来てサルによる落葉を採食し始めた。さらにサルが採食を終了し、採食樹から去った後も、シカは樹下に残って採食を続けていた。
他方、カンコノキのようにサルの樹上採食頭数が増加しても、シカはほとんど樹下採食をしなかった樹種もあった(図6)。 カンコノキはトゲがあるためにシカが採食しなかったのかもしれない。
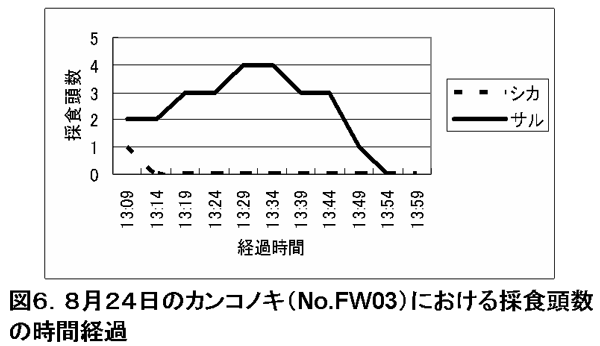
図6. 8月24日のカンコノキ(No.FW03)における採食頭数の時間経
なおアコウでは、サルが以前に樹上採食によって落とした果実を、サルがその場にいない状況でシカが採食していた。このことから、今回はシカがアコウを訪れた時に、サルが樹上採食していなかったが、もしサルが樹上で採食している状況であれば、その場でシカも樹下採食をしたと考えられる。また、アコウの実は、サルの樹上採食時に大量に落とされるため、時間が経過した後でもシカがその実を利用することができたと考えられる。 今回の調査によって、樹種によって若干の違いはあるが、特定の採食樹において、サル、シカ、の順で訪れ採食し、サル、シカの順で去っていくということが明らかになった。
(住栄 貴恵)
図7は、2006年8月24日、サルE群とシカのダニーの遊動の様子を示している。E群とダニーの移動ルートが途中からおよそ600メートルにわたり重なっている。つまり、ダニーは単に樹下食いのために一時的に集まってくるだけでなくE群を追随していることが分かった。

図7. 2006年8月24日、サルE群とシカのダニーの遊動
図8、および図9はその追随の間にみられた樹下採食におけるサルとシカの採食頭数の時間推移を示している。この図からも採食樹には先にサルがやってきて、その後シカがその樹下にやってきて採食する。そしてサルが採食を終え立ち去った後も、シカは採食を続け、しばらくしてからその場を立ち去ることが分かる。
(下中 麻奈美)
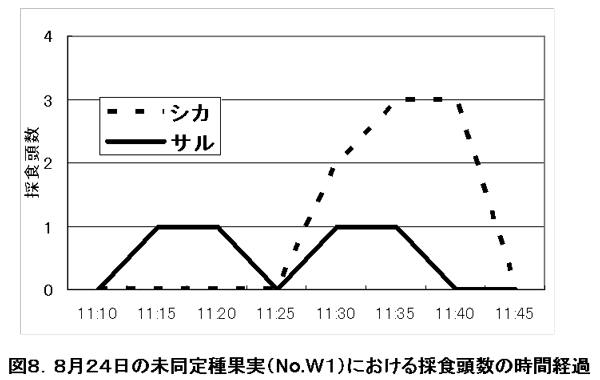
図8. 8月24日未同定果実(No.W1)における採食頭数の時間経
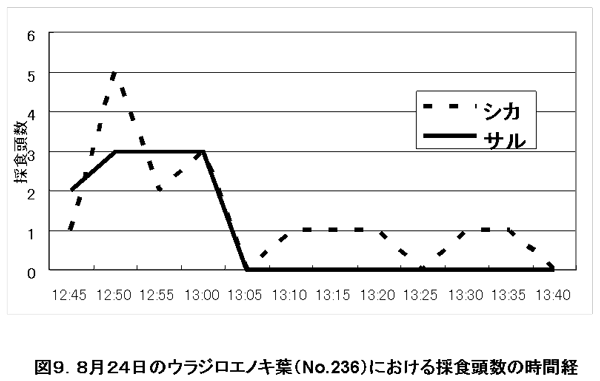
図9. 8月24日のウラジロエノキ葉(No.236)における採食頭数の時間経
ケース1(8月24日):ダニーはE群を追いかけていた。ウラジロエノキ樹下で採食中に立ち去るE群を突然追わなくなり、ウラジロエノキ付近で採食・反芻を続けた。50分後E群は再び採食のために戻ってきた。
ケース2(8月27日):ダニーはアコウ(c25)樹下付近に1.5時間滞在し、移動後再びアコウに戻り滞在していた。この間E群はこの付近にはいなかった。なお、これらの木は普段サルがよく利用する採食ポイントであり、シカも好んで採食するが高い位置に枝があるため自力では採食できない。
以上より、ダニーは自分のホームレンジ内でサルがよく利用する採食ポイントで待ち伏せし、サルを追うよりもより効率的に採食しようとしているのではないだろうか。
樹下でのサルの採食時のクーコールには1頭、樹上でそれには4頭が反応(音源注視・移動・耳を動かす)した。音声に対するシカの反応には個体差があるが、樹上での音声には特に反応を示した。
以上よりシカはサルの音声、葉音を採食場所の指標として追従・待ち伏せをしていると思われる。
(増山 遥)
図10は排泄からの経過時間とその時点での糞の分解度を示している。分解度0はまったく形が崩れていない状態を指し、値が上がるほど分解の程度が高く、分解度4が完全な消失を指す。
サルの糞の分解、消失には二つの原因があげられる。一つはシカによる採食。もう一つは糞虫によるものである。前者の場合は、糞は完全に道路上から消失してしまう。後者は糞の残骸が少なからず残る。このことにより、二つは区別可能である。図10から5時間以内に40%が完全に分解、消失されており、これらはシカによる採食と考えられる。つまり、シカはサルが排便してから、あまり時間をおかずに、それを採食していると考えられる。
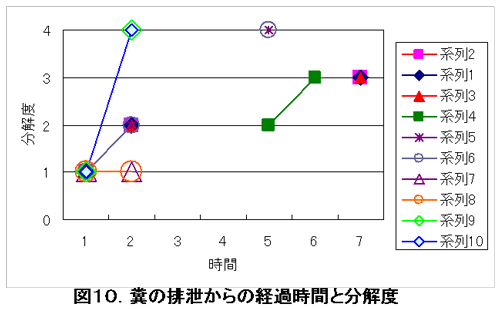
図10. 糞の排泄からの時間経過と分解度
上記のような採食に関する種間関係以外にもヤクシマザルとヤクシカの間にいくつかの種間の社会的交渉が観察された。
8月23日、サルE群がグルーミング中にシカ(3尖オス)が接近していった。すると、木に上っていた1頭のサルが下りてきて、そのシカの首のあたりに跨り乗った。そして、シカに対しグルーミングを行った。その間シカは暴れることなくじっとしていた(図11)。

図11. サルによるシカの背乗りグルーミング
グルーミングしてもらうことはシカにとってはダニやシラミを食べてもらえるという利益を得られ、サルにとってもそれらの採食という意味では利益であり、相利共生的関係であるとも考えられるのかもしれない。
あるサルがグルーミング中に、背後からシカが近距離まで近づいた。しかしサルはそれに全く気づいておらず、振り返ると突然シカがいたため、驚いてそのシカに対し威嚇した。しかしこの時シカはサルに対して攻撃的な行動は示さなかった。
以上より、採食行動に関しては、サルとシカの間にはシカにとってのみ利益のある片利共生関係が認められた。そ他にもグルーミングや背乗り行動など直接的な種間の社会交渉もみられた。半山でのサルとシカはこうした種間関係のもと緊密になっており、彼らが同時に観察されることが多くなったと、この調査から考えられる。
(下中 麻奈美)
(増山 遥)
京都大学野生動物研究センター>屋久島フィールドワーク講座 >第8回・2006年の活動-サル班-報告書
このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹