| 京都大学野生動物研究センター > 屋久島フィールドワーク講座 > 第6回 2004年の活動-シカ班-報告書 | 最終更新日:2004年12月17日 |
| 概要 | 人と自然班 | シカ班 | 植物班 | 博物館班 | サル班 | 公開講座 | スタッフ |
| 報告書 | 写真集 | 感想 |
参加者:代口麻衣美・石場圭太・船田恵子・木村桃子・西村耕野
講師:揚妻直樹・日野貴文
通常、森林内には細い木の密度が非常に高く、太い木の密度が低いというL字型構造を示すことが多い。しかし、シカが高密度に生息している地域では、シカの採食圧によって森林構造が著しく改変されると考えられている。例えば、北海道の洞爺湖ではシカ密度が1k㎡あたり30頭を越えてから、ほんの数年で細い木がほとんどなくなってしまったことが報告されている。このようにシカ密度が高い地域では森林構造のバランスが崩れた状態になるようである。
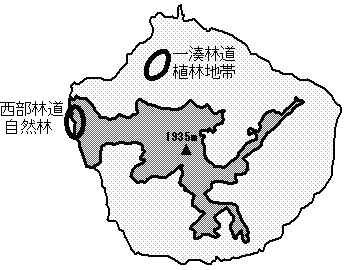
屋久島の西部林道周辺の自然林(図1)には1k㎡あたり60~80頭ものシカが生息しており、そこは日本の中でも非常にシカ密度が高い地域といえる。そこで、我々はこの自然林でも森林構造が改変されているのかを検証することにした。そのため、この地域の森林構造とシカの採食圧、生息密度、行動範囲、行動、食性を調査した。また、屋久島には自然林が伐採され、スギの植林が広がっている地域も多い。そこで、こうした植林地帯(図2)においても森林構造とシカの採食圧・生息密度を調査した。
図1 調査地。
楕円で囲った部分が一連の調査を行った場所。西部林道周辺の自然林は約50年生の広葉樹林と一次林で構成されている。一方、一湊林道周辺は約半分がスギ植林地に占められ、残りが若い二次林、古い二次林、一次林などの広葉樹林で構成されている。濃く色がついている部分は世界遺産地域を示す。
自然林と植林地帯でのシカ密度を把握するために、2004年8月17日と18日に屋久島西部の自然林(西部林道の半山地域)と森林の半分がスギ植林に置き換わった北部の植林地帯(一湊林道)で調査を行った(図1)。西部林道に約2.2km、一湊林道には約2.4kmの調査路を設定し、日中にそこをゆっくり歩いてシカの発見に努めた。シカを発見した場合には、その場所と時間、シカの性別と年齢を記録した。
自然林ではオス3頭、メス16頭、当歳児5頭の合計24頭を発見した。一方、植林地帯では性年齢不明のシカ1頭を発見するに留まった。調査路1km当たりのシカ発見率は自然林では11.1頭、植林地帯では0.4頭となり、自然林では植林地帯に比べ28倍もシカが多いことが解った。
シカが高密度で生息することで森林構造が変化するのかを明らかにするために、シカ密度の高い西部の自然林(西部林道の半山地域)において植生調査を行った(図1)。この地域では1989年に植生調査が行われていたので、その時と同じプロットで調査を行い、2004年までの15年間における森林構造の変化を検討することにした。
調査は2004年8月17日に行った。1989年に尾根と谷にそれぞれ設定した5×25mのコドラートについて、胸高直径5㎝以上の樹木(幹)を対象に、1989年調査当時の幹番号、種名を記録し、胸高周囲長を計測した(πで割り胸高直径とする)。さらに、幹に対するシカの採食痕や角こすり痕の有無を調べた。痕を発見した場合は樹皮が失われた部分が幹周りの何割に達していたか、その痕の新鮮さ・痕の回復の程度も記録した。また、胸高直径5cm未満で樹高が40cm以上の樹木については、種名・個体数とシカの幹の採食痕・角こすり痕の有無と新鮮さを記録した。
図2は2004年の自然林の構造を表している。この結果から2004年の自然林では細い幹の密度が圧倒的に多く、太い幹ほど密度が低いL字型の構造を示していることが解った。なお、最も小さい木が属する胸高直径5㎝未満の階級には高さ40cm未満の稚樹が含まれていないので、実際にはこの階級にずっと多くの個体があったといえる。
シカによる幹への加害としては角こすり痕だけが見られ、しかも痕はすべて何年か前につけられたと思われる古いもののみであった。調査した幹226本の中で角こすり痕が見られたのは23本で、その割合は10.2%であった。角こすりをされていた幹のほとんどは胸高直径5㎝未満であった。
次に1989年と2004年の植生構造の比較を行った。1989年には胸高直径5㎝未満の個体を調査していなかったため、胸高直径5㎝以上の幹数をのみを比べた(図3)。その結果、この15年間で各階級の幹数にほとんど変化がないことが解った。また、1989年に調査された幹のうち、この15年間で枯死したものは21本であったのに対し、この15年間で新しく胸高直径が5cm以上に成長してきた幹は22本あった。従って、枯死木と更新木のバランスが保たれていたといえる。これらのことは、この自然林では森林の更新はうまく行われており、シカの採食圧による森林構造の劇的な変化は起きていないことを示唆している。
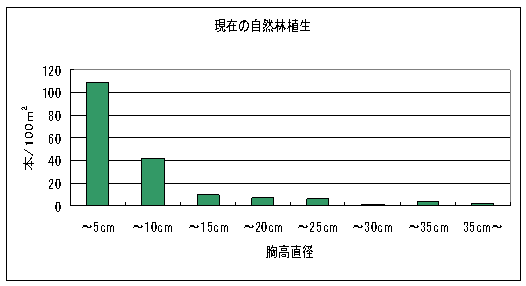
図2 2004年の自然林の構造。胸高直径の階級は5㎝きざみになっている。縦軸は100m2あたりに換算した幹本数を表す。
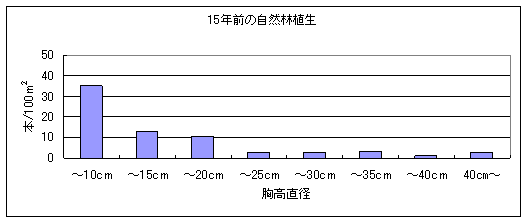
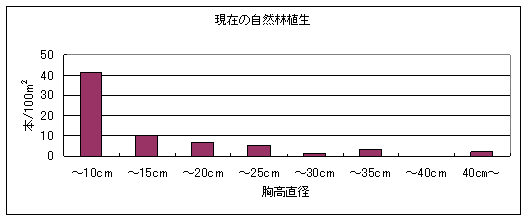
図3 1989年(上)と2004年(下)の胸高直径クラスと密度の関係。
屋久島の中標高以下にはスギ植林と二次林などの広葉樹林がモザイク状に混合した植林地帯が広がっている。ここでは屋久島で見られる典型的な森林として、この植林地帯の構造を把握を行い、自然林との比較を試みた。2004年8月18日に、一湊林道周辺(図1)の植林地帯の中にある広葉樹二次林とスギ植林にそれぞれ10×10mのコドラート設置した。そして、自然林での植生調査と同じ要領で、胸高直径5㎝以上の木(幹)について種名、胸高周囲長を記録し、シカによる幹に対する採食痕や角こすり痕を調べた。また、胸高直径5cm未満で樹高が40cm以上で木についても同様に種名、個体数、シカの採食痕・角こすり痕を調べた。
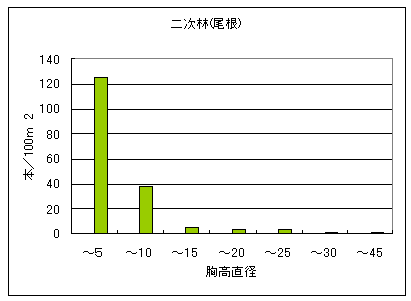
図4に植林地帯の中の二次林(右)とスギ植林(左)の構造を示した。二次林では細い幹の密度が圧倒的に多く、太い幹ほど密度が低いL字型の構造を示していた。なお、このコドラートで見られた樹種はのうちの53%は自然林では見られなかったものであった。総調査本数(177本)に対する樹皮に対する採食痕があった割合は3%、角とぎ割合は3%であった。ただし、これらの傷跡は全て古いものだった。
一方、スギ植林では胸高直径20cm以上のものはスギだけで、成長した広葉樹は見られなかった。これはスギが日光を遮り、広葉樹の生長を阻害しているためだと考えられた。植林地で見られた樹種の55%は自然林では見られなかったものだった。スギ植林内には樹皮採食、角とぎ共に見られなかった。
植林地帯におけるシカの幹に対する傷痕は自然林と比較して少なかった。これはシカの生息密度と関係が
あると考えられる。自然林では植林地帯の28倍も多くシカが発見されていた。これほどまで植林地帯でシカ密度が低いのは、そこが自然林と比べ生産性が低く、シカにとっての資源が少ないということを示唆している。
 |
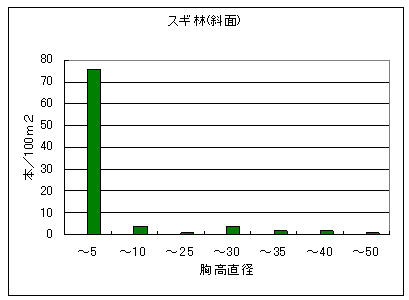 |
図4 植林地帯の中の二次林(左)とスギ植林(右)の構造。横軸は5cmきざみの直径階級を、縦軸は100m2あたりの本数を示す。
 |
 |
| 写真1 スギ植林での植生調査の様子 | 写真2 自然林での植生調査の様子 |
自然林ではシカによる著しい植生改変が確認されなかった。この理由を考える上で、自然林内でのシカの行動、特に食性を明らかにすることは重要である。そこで、西部の自然林(半山地域:図1)において、2004年8月19日と20日にシカの行動観察を行い、行動内容と食物を調べることにした。
観察方法には個体追跡法を用いた。森林内で視界に入ったシカの中から1頭を選び、その行動をできるだけ長く観察し、各2分間の中で最も多く行っていた行動を記録していった。なお、シカの行動は以下の9つのカテゴリーに分けた。また、観察中にシカが採食した場合には、その食物を記録した。追跡個体を見失った場合には別の個体を探し、新たな追跡個体として観察を行った。
行動カテゴリー
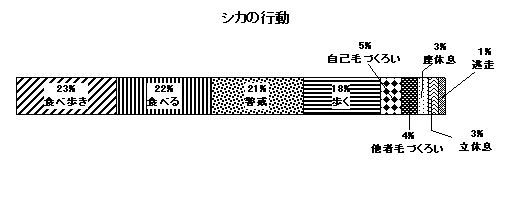
図5 総観察時間に占める各行動カテゴリーを行っていた割合
シカの成熟オス・メス、未成熟個体を対象に全部で560分間(280データ)の行動を記録した。総観察時間に対してシカが行った各行動カテゴリーの時間割合を図5に示す。ここから、シカの行動の約半分(45%:食べ歩き+食べる)が採食に費やされていることが解る。警戒については観察者がいたことで割合が増えた可能性が高く、通常はもっと少ないと考えられた。
次に、シカが採食していた食物の割合を図6に示す。彼らは林床に落ちた葉・果実、樹皮、シダ植物などを採食していた。なお、図6の「落ちていたもの」の中の「不明」とは、シカが林床から何か拾いあげて食べたことは確認できたが、食物の種類が特定できなかったものをまとめた。また、「落ち葉」にはアカメガシワ、イイギリ、イヌガシ、クマミノズキ、ヒメユズリハなどの広葉樹の緑色のもの、紅葉したもの、茶色く枯れたものが含まれていた。シカは植物の生きている部分(生きていた植物)も採食するものの、林床に落ちている葉や果実など(落ちていたもの:リター)を採食する割合が圧倒的に高く、9割を占めていた。
この自然林でシカの採食による森林への影響が低く抑えられていたのは、シカが植物の生きている部分をあまり採食しないためと考えられた。ただし、シカがリターを多く採食する理由についてはよく解らない。シカは単にリターに対する嗜好性が高いのか、本当は生きた植物に嗜好性があるが、木の高いところにあって利用できないのかは、今後明らかにしていくべき課題といえよう。
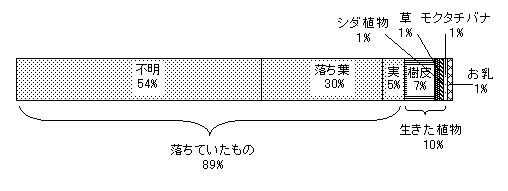
図6 シカが採食した食物の割合
 |
 |
| 写真3 シカの行動観察をしている様子 | 写真4 行動観察中に現れたメスシカ |
自然林で植生構造の著しい改変が見られないのは、シカの移動範囲が広く、数km以上離れた場所にある果樹園などで質の高い食物を得ていたからかもしれない。そこで自然林(西部の半山地域:図1)に生息するシカの移動範囲がどの程度なのかを把握するために、ラジオテレメトリー法による調査を行った。
この地域の2頭の成熟したメスシカ(さんちゃん、きゅーちゃん)に付いている発信機の電波を30分以内に2~3地点から指向性のある八木アンテナで受信し、シカがいる方向を割り出した。そして、2地点から方法を割り出した場合にはそれらの方向が作る交点を、3地点から方向を割り出した場合には、それらの方向が形づくった三角形の重心をシカの位置とした。このシカの位置の測定を連続的に繰り返し、位置を結んでいくことでシカの移動距離・移動範囲を推定した。この調査は2004年8月20日の午前9時47分から午後4時7分まで約7時間行った。
調査の結果、さんちゃんの移動距離は1704mで移動ルートの最大幅は387m、きゅーちゃんの移動距離は515mで移動ルートの最大幅は333mであった(図7)。日中のみの調査なので確実なことは言えないが、移動ルートの最大幅(333~387m)を見る限り、シカは半山地域の中でウロウロしており、数kmも離れた果樹園に移動するほどは動いていないように思われた。
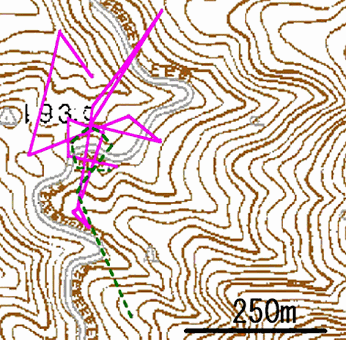
図7 メスシカのさんちゃん(実線)と、きゅうちゃん(破線)の7時間の移動ルート

写真5 八木アンテナで電波の方向を探っている様子
屋久島西部の自然林ではシカが高密度に生息しているにも関わらず、この15年の間、著しい植生の改変が起きていなかった。そして、樹木の死亡数と更新数もつりあっていた。このことはシカの採食や角とぎによって森のバランスが崩されていなかったことを示唆している。その理由として、稚樹の採食や角とぎやなど、森林に対して悪影響を及ぼす行動が他地域より少ない可能性が考えられた。シカの行動観察から、シカは林床に「落ちていたもの:リター」を多く食べ、植物の生きた部分をあまり食べていなかった。また、角研ぎをされていた木の割合も他のシカ高密度地域よりは少ないようで、しかも発見できた傷はすべて古いものばかりだった。このためシカによる植生への悪影響が低く抑えられていたと考えられる。
シカの行動観察中に、近くに来ていたサルが木の上から落とした果実をシカが採食しているのが見られた。シカの植生への影響が低いのは、サルが木の上にある質の高い食物をシカに供給しているからかもしれない。しかし、宮城県の金華山にはシカとサルが高密度で生息しているが、シカによる著しい植生改変が報告されている。このことから、サルがいたからといって必ずしも植生への影響が緩和されるわけではないのかもしれない。
自然林(西部林道)と植林地帯(一湊林道)ではシカの生息密度が大きく異なっていた。植林地では植林後20年もすると動物にとっての食物生産性が非常に低下することが解っている。一湊林道周辺のスギ植林は30~40年生のものが多かったので、そこでは食物生産性がかなり低下していたと推測できる。シカの生息密度が低かったのはその影響によるものと言えるかもしれない。ただし、一湊林道周辺ではスギが植林される前からシカが少なかった可能性、植林とは別の理由でシカが少なくなった可能性もある。従って、シカ密度の異なる様々な地域の特徴を調査し、シカ密度と環境の関係を明らかにしていく必要がある。
京都大学野生動物研究センター > 屋久島フィールドワーク講座 > 第6回 2004年の活動-シカ班-報告書
このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹