| 京都大学野生動物研究センター > 屋久島フィールドワーク講座 > 第6回・2004年の活動−植物班−報告書 | 最終更新日:2004年12月20日 |
| 概要 | 人と自然班 | シカ班 | 植物班 | 博物館班 | サル班 | 公開講座 | スタッフ |
| 報告書 | 写真集 | 感想 |
参加者:飯田佳子・小島正行・常木静河・中島淑子
講師: 村上哲明・篠原 渉
今回、植物班では屋久島の高地で小型化したと考えられている植物種の調査を行った。屋久島は、固有植物種(ここにしか分布していない種)が非常に多いことが重要な特徴の一つである。特に屋久島の高地には多数の固有種や固有変種が生育しているが、これらは低地に生育する近縁種と比較して植物体が小型化したものが多い。しかし、これらの植物がなぜ小型化しているのか、どのような環境に適応した結果なのか、その要因はよく分かっていない。また一概に小型化と言っても、どのように小型化しているのか、すなわち、すべての器官が同じ割合で小型化しているのか、それとも器官ごとに小型化の程度に違いがあるのか、さらに植物体を構成している細胞数が減ることによって小型化しているのか、それとも個々の細胞が小さくなって小型化しているのかということなども詳細に調べられたことがない。そこで本講座では、これら屋久島の高地で小型化したと考えられる植物種が、低地に生育する近縁種と比較してどのように小型化しているのかを、植物体レベル及び細胞レベルでの比較から明らかにしようと試みた。さらに被子植物とシダ植物という系統的にかけ離れた分類群の小型化する機構を比較することにより、小型化の様式に差がないか、逆に遠縁の植物群間でも共通して起こっていること(適応や発生上の制約などによる可能性がある)がないかを調べた。
木本としてオオヤクシマシャクナゲとヤクシマシャクナゲ、草本としてコナスビとヒメコナスビ、シダ植物としてシシガシラとヒメシシガシラの3組の植物群を対象に調査をおこなった。なおこの3組において前者がより標高の低い場所に生育しており、後者がより標高の高いところに生育している。
ヤクシマシャクナゲ(図1、2)は屋久島の固有変種で標高1000m以上の高地に生育しており、高度が上がるにつれて葉のサイズが小さく、葉の裏面の毛が密になる傾向がみられることが報告されている。林内に生育しているものはより高所の笹原に生育しているものと比較して、葉のサイズが大きく、前者をオオヤクシマシャクナゲ(Rhododendoron
yakusimanum var. intermedium)、後者をヤクシマシャクナゲ(Rhododendoron
yakusimanum var. yakusimanum)として区別することもある.しかし、両者は連続的に生育し、どの形態形質においても連続的に変異が見られる両者を明確に区別することは困難であると多くの研究者は考えてきた。
今回の講座では淀川登山口(標高約1400m)から小花之江河(標高約1600m)にかけての登山道沿いに生育するヤクシマシャクナゲ類について高度の異なる複数の地点から採集を行い、葉の大きさや毛の量が高度に応じてどのように変異するかを調べた。具体的には、海抜高度を高度計により測定しながら様々な高度で葉を採集し、地図上に採集地点を記録した。また採取した葉を宿舎に持ち帰り、葉の幅と長さ、および葉の裏面の毛の量を測定した。毛の量は、少ない、やや多い、多いの3段階で評価した。
ヒメコナスビ(Lysimachia japonica var. minutissima)は屋久島の高地(1500m以上)にのみ生育するコナスビ(Lysimachia japonica var. japonica)(図3)の変種であり、コナスビと比較して植物体が小さい状態で花をつけるのが特徴である。また屋久島の低地にはコナスビが普通に生育しているが、両者の形態を詳細に比較した研究はこれまで報告されていない。
コナスビについては8月17日に湯泊林道沿いの集団(標高約400m、以下Y集団)から19個体、小楊枝林道沿いの集団(標高約80m、以下K集団)から13個体、一方、ヒメコナスビについては8月18日に小花之江河の集団(標高約1600m、以下YDK集団)から12個体採取した。採集は個体を根ごと掘り起して、付着した泥を水で洗い流した後、個体ごとに固有の番号をつけビニール袋に入れて持ち帰った。今回、採取した全ての個体は結実しており、成熟した個体であると考えられる。採取したすべての個体について最大葉、茎、根の長さを測定した。また、全重量に対するそれぞれの器官の乾燥重量の比を各集団で比較するために、採集した全個体を京都大学屋久島ステーションの乾燥機で完全に乾燥させた後、根・茎・葉および生殖器官に分解し、それぞれの器官の乾燥重量を測定した。さらに葉の表皮細胞の大きさを調べるため、スンプ法(Suzuki's
Universal Micro-Printing Method)により葉の裏面の表皮細胞群のレプリカをとり、それを光学顕微鏡により観察した。
ヒメシシガシラは屋久島の1000m以上の高地にのみ産するシシガシラ(Blechnum
niponicum)の一型で典型的なシシガシラと比較して植物体が小さい状態で胞子葉をつける(図4左)。また屋久島の低地には普通のシシガシラ(図4右)も生育している。
今回は、シシガシラについては8月20日に荒川分れ(標高約720m、以下AW集団)にて、またヒメシシガシラについては8月19日に淀川小屋から小花之江河にいたる登山道沿い(標高約1600m、以下YDS集団)において採集を行った。シシガシラは胞子葉をつけている11個体を、またヒメシシガシラについては同じく胞子葉をつけている8個体を採集した。採集はコナスビ類と同様に根から掘り起こし、付着した水で土を洗い流した後、ビニール袋に入れ持ち帰った。それぞれの個体の最大の栄養葉と胞子葉の幅および長さを測定した。またスンプ法を用いて葉の裏面の表皮細胞を観察した。
|
|
| 図1 ヤクシマシャクナゲの花 | 図2 ヤクシマシャクナゲ(淀川小屋付近) |
|
|
| 図3 コナスビ (湯泊林道にて) | |
| 図4 ヒメシシガラシ(左)・シシガラシ(右) |
ヤクシマシャクナゲは葉の裏面の毛の量と海抜高度の間に正の相関が見られ、高度が高くなるにつれて葉の裏面の毛の量も多くなっていた(図5)。
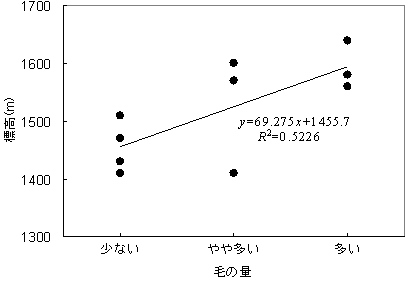
図5 ヤクシマシャクナゲの葉裏面の毛の量と生育していた標高の関係
葉長と茎の長さの比較ではより高地のYDK集団の個体はより低地のY集団やK集団の個体と比較して葉長と茎の長さ共に小さくなっていた(図6)。
乾燥重量比では、YDK集団は全体に対する葉の占める割合が他の集団と比較して小さく、逆に生殖器官の占める割合はYDK集団では大きく、Y集団及びK集団では小さかった(図7)。このことからヒメコナスビでは器官によって小型化した割合が異なっており、葉が特に小型化し、次に茎と根が小型化しており、それに比べて生殖器官はあまり小型化していないと言える。
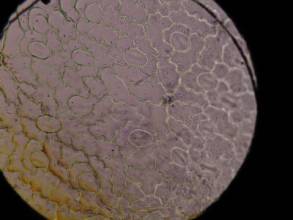
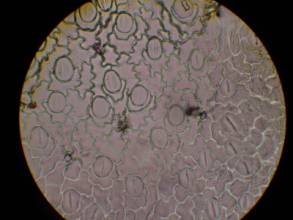
表皮細胞の観察から、YDK集団の個体はY集団の個体と比較して表皮細胞のサイズが小さくなっていることがわかった(図8)。
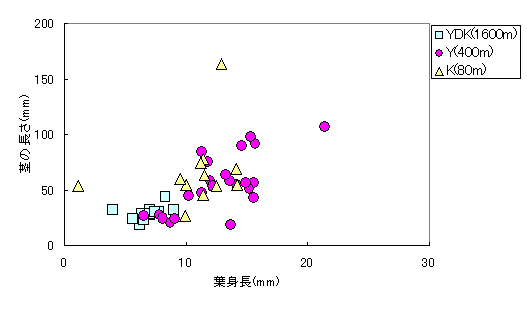
図6 コナスビ類の成熟個体の葉身長と茎の長さ
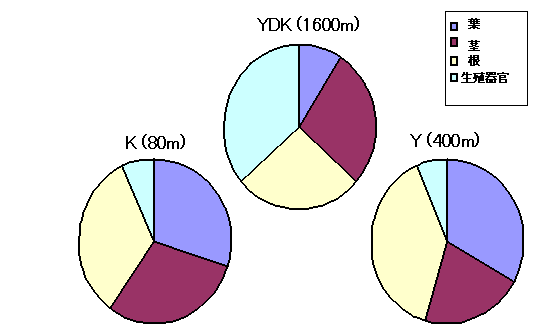
図7 コナスビ類の葉・茎・根の乾重量比
| Y (400m) | YDK (1600m) | |
 |
 |
|
図8 コナスビ類の葉の表皮細胞の大きさの比較
ヒメコナスビでは葉が特に著しく小型化していたことから、ヒメコナスビの小型化に適応的意義があるとするならば、葉の大きさに特に強く作用する環境要因に対する適応であろうと考えられる。風による物理的損傷や風による乾燥などは、このように特に葉を小型化させる環境要因として理解しやすい。
一方、栄養器官が著しく小型化しているのに対して生殖器官がそれほど小型化していない理由として、生殖器官は次世代を残すための重要な器官でありに、小型化することが適応的ではなかったためではないかと考えられる。花のサイズは訪花昆虫に対応しているはずで、高地でも同じあるいは同じような大きさの昆虫が来るのであれば、花のサイズも小型化させることが適応的でないことは容易に想像できる。また、種子のサイズについても、小型化してしまえば貯蔵できる栄養分が少なくなって、やはり適応的でないことが予想される。
表皮細胞の比較からヒメコナスビでは個々の細胞の小型化が植物体の小型化の一因であると考えられる。ヒメコナスビの葉の細胞数がコナスビと比較して変らないかどうかについては、葉全体の表皮細胞数の比較を行なったうえで再検討する必要があるであろう。
葉の細胞が小型化するとすれば、葉と相同な器官である花も一緒に小型するのが自然である。今回の調査では、特にヒメコナスビの花が完全に終わっていたので、花の大きさは比較をすることができなかった。しかし、果実の大きさを比較すると両者でそれほど違わなかったことから、花も葉ほどには小型化していないと予想される。図鑑の記載などを見ても、花の大きさはそれほど変わらないようである。とすれば、細胞の小型化は栄養器官である葉だけで起こっているのかもしれない。将来的に、花(例えば花弁や顎)についても細胞の大きさを比較してみると興味深いと考えられる。
低地のAW集団と高地のYDS集団の栄養葉と胞子葉の大きさを比較したところ、YDS集団では、栄養葉、胞子葉共にAW集団と比較して小型化している傾向が見られた。しかしながら栄養葉と胞子葉の小型化している程度には違いが見られ、胞子葉は栄養葉ほどは小型化していなかった。さらに、YDS集団の個体では、栄養葉の長さにかかわらず胞子葉の長さはより一定である傾向も見られた(図9)。
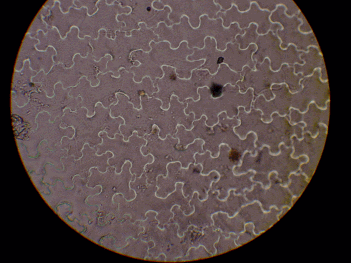
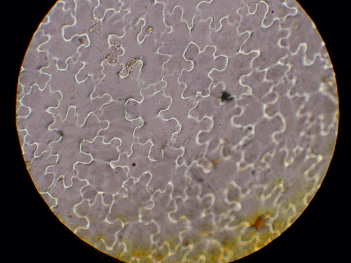
スンプ法による葉の表皮細胞の観察からAW集団とYDS集団では表皮細胞の大きさに差異はみられなかった(図10)。
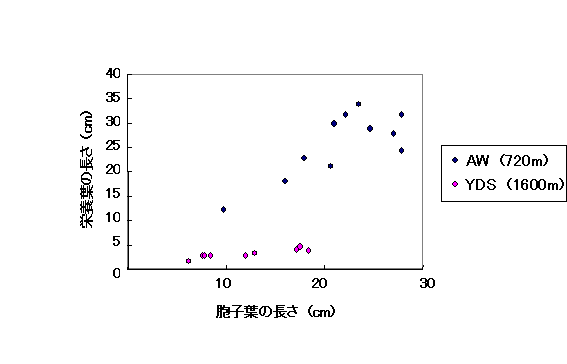
図9 シシガラシ類の胞子葉と栄養葉の長さ
| AW (720m) | YDS (1600m) | |
 |
 |
|
図10 シシガラシ類の表皮細胞の大きさの比較
シシガシラと比較してヒメシシガシラの胞子葉が栄養葉ほど小さくなっていなかった理由として、種子植物のヒメコナスビで見られたのと同様、胞子葉は次世代の胞子をつくる重要な生殖器官であるために、胞子葉が小さくなることは適応的でないためではないかと考えられる。さらに、胞子を効率よく散布するためには最低限ある高さが必要なのかもしれない。仮に強風などでより傷害を受けやすくなってでもある。胞子葉は、栄養葉よりもより短期間(胞子を散布する期間だけ)立った状態で存在すればよいことも、相対的に小型化していないことに関わっている可能性がある。
またヒメシシガシラとシシガシラでは細胞の大きさに差異が見られなかったことから、ヒメシシガシラではシシガシラよりも細胞数が減少することによって小型化していると考えられる。栄養葉も胞子葉も、ともに茎頂で全く同様の過程で発生する。今回、細胞の大きさを調べたのは栄養葉についてだけであるが、胞子葉の細胞がどのようになっているかも今後に残された興味深い課題である。
ヒメコナスビとヒメシシガシラ共に葉(栄養葉)が小型化しており、生殖器官は葉ほど小型化していないという共通の傾向がみられたことは大変興味深い。このことから生殖器官を小型化することは、適応的ではない場合が多い可能性がある。今後、他の小型化した植物群でも、近縁種との同様の比較をしてみる必要があるだろう。
また細胞の大きさではヒメコナスビではコナスビに対して小さくなっていたのに対して、ヒメシシガシラではシシガシラに対して細胞の大きさに違いはみられなかったことから、両者の間では葉を小型化するメカニズムが異なっていると考えられる。系統的に離れた2種が異なる様式により葉を小型化させていたことは興味深い。屋久島の高地では葉(特に栄養葉)を小型化することに強風による物理的破壊や乾燥から逃れられるなどの点で適応的意義がある可能性が高いと考えられる。とすれば、それぞれの植物群ごとに他の形質に悪影響がでにくい、しかも少数の遺伝子(最初はたった一つの突然変異から始まるはず)の変化で効率よく必要な器官が小型するようになると考えられる。このような様式が被子植物とシダ植物で異なっていたとしても不思議ではない。他の被子植物とシダ植物でもヒメコナスビやヒメシシガシラで見られたのと同様の変化が見られるのかどうかを調べる必要があるだろう。
渓流沿い植物とその近縁種の葉の細胞の大きさ比較した研究(Kato and Imaichi,1992
Usukura et al.,1994)では被子植物では細胞の大きさに変化が見られず、逆にシダ植物では細胞が小型化していた。今回の屋久島の高地での小型化植物では被子植物では細胞が小さくなっており、シダ植物では細胞の大きさが変化しないため、渓流沿い植物の葉の狭小化とはその小型化のメカニズムが被子植物とシダで逆になったことは非常おもしろい。一般的に植物は最初に細胞増殖を行う細胞増殖期があり、その後、個々の細胞が伸張する細胞拡大期がある。加藤(1999)は渓流沿い植物の葉の狭小化現象について、個体発生の形成開始・終了時期などのタイミングのずれや成長速度の変化が形態を変化させる異時性が関わっている可能性があるとしているが、ヒメコナスビとヒメシシガシラについても、前者は細胞拡大期が短くなり、後者は細胞増殖期が短くなるといった異時性がそれぞれの形態の形成に関与している可能性が考えられる。葉の発生の段階ごとの比較観察およびそれらを司っている遺伝子レベルの比較解析によって、これら異時性の問題についてさらに理解を深めることが必要である。
またさらに多くの屋久島高地で小型化する植物とその近縁種の解析を行うことにより、なぜ小型化するのかという問題について理解が深まることが期待される。今回の調査では、ちょうど台風が接近する中、私たちは高地に上り、ヒメコナスビとヒメシシガシラを採集した。そして、特に周りの開けた小花之江川では驚くほどの強風にあった。大きな葉や高い茎の植物がこのような環境下で適応的でないことは実感できた。実際、ヒメコナスビでは特に葉が一番著しく小型化していたので、屋久島における小型化植物の進化が強風に対する適応である可能性は十分ある。ただ、土壌や温度など他の環境要因も複合的に関与している可能性はまだ十分残っている。今後、なぜ(どのような適応的意義があって)多くの植物種が小型化しているのかについても解明していく必要があるだろう。
Kato
M. and R. Imaichi. 1992. Leaf anatomy of tropical fern rheophytes with its evolutionary and ecological implications. Can. J. Bot. 70
165-174.
Usukura
M.
R. Imaichi and M. Kato. 1994. Leaf morphology of a facultative rehophyte
Farfugium japonicum var. luchuense (Compositae). J. Plant Res.
107
263-267.
加藤雅啓.1999.植物の進化形態学.東京大学出版会
京都大学野生動物研究センター > 屋久島フィールドワーク講座 > 第6回・2004年の活動−植物班-報告書
このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹