| 京都大学野生動物研究センター>屋久島フィールドワーク講座 >第7回・2005年の活動-シダ班-報告書 |
| 概要 | 人と自然班 | サル班 | シダ班 | ヤモリ班 | 公開講座 | スタッフ |
| 報告書 | 感想文 |
参加者:高木菜穂子・浜 一朗・秀島瑠満子
講師:村上哲明・石川 寛・篠原 渉
シダ班では屋久島に生育するベニシダ類を材料として、無配生殖型と有性生殖型が屋久島でどのように共存しているのか、生育環境を分化させるなどして異なる生殖型が共存しているのかを調べた。同時に、異なる生殖型間、および同じ生殖型内の形態の違いについても観察をおこなった。
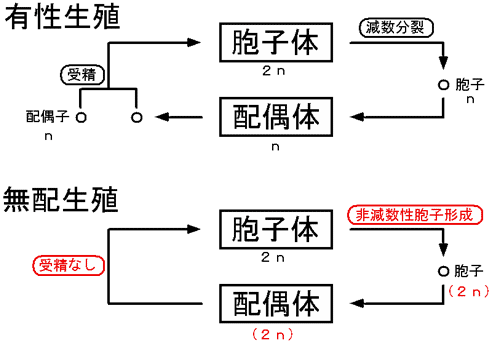
まず、シダ植物の無配生殖について説明する。多くのシダ植物は、生活環の中に減数分裂と受精を含む「有性生殖」をおこなう。薄嚢シダ類の大多数の種では、胞子体(普段、シダ植物として見ているもの)の上(主に葉の裏)にできる胞子嚢の中で、1個の胞原細胞から4回の体細胞分裂で16個の胞子母細胞がつくられ、母細胞が減数分裂してそれぞれ4個の胞子となり、胞子嚢あたり64個の、染色体数の半減した胞子が形成される。胞子は発芽して配偶体(前葉体)となり、造卵器と造精器をつくる。卵と精子が受精すると、親胞子体と同じ染色体数の次世代の胞子体が発生する。これに対して、「無配生殖」では減数と受精の過程が省略されている。無配生殖型の胞子嚢の中では、染色体数の倍加した8個の胞子母細胞がつくられ、それらが減数分裂することで、結果的に胞子体と同じ染色体数の32個の胞子ができる(図1、2)。
図1 シダ植物における有性生殖種と無配生殖種の生活環
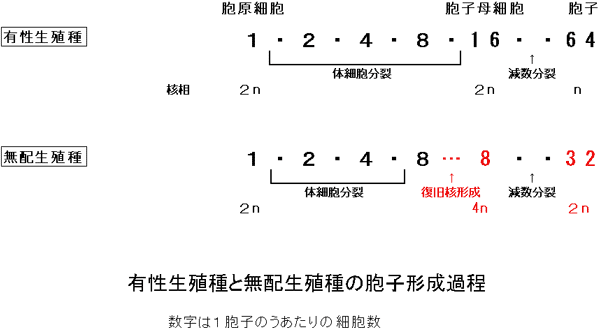
図2 シダ植物における有性生殖種と無配生殖種の胞子形成過程
胞子は発芽して非減数性の配偶体となるが、卵と精子が受精することなく、前葉体のクッション部(中央部の細胞層の厚いところ)から次世代の胞子体が発生する。つまり無配生殖とは一種の無性生殖であり、無配生殖をおこなう個体から発生した次世代胞子体は、基本的に親個体と遺伝的組成がまったく同一のクローンとなる。
次世代に残す子孫の数が一定という仮定のもとでは、無配生殖は有性生殖に比べて次世代により自分自身の遺伝子を二倍多く残せるため、無配生殖は有性生殖より増殖の上で有利な生殖様式であると考えられている。また無配生殖では、有性生殖にみられる卵と精子の接合という大きな不確定要素を含むステップが存在しないため、より効率的に子孫を残せるとも考えられている。
つまりシダ植物の無配生殖型と有性生殖型の間に、生育環境の分化など競争を緩和する要因が存在しなければ、無配生殖型の方がその増殖力の高さゆえに優占し、有性生殖型が無配生殖型と混生することはきわめて困難であると考えられるのである。実際に日本のベニシダ類において、屋久島や八丈島など、限られた地域を除けば、無配生殖型のほうが圧倒的に多い。したがって、これまで屋久島において報告されているベニシダ類の無配生殖型と有性生殖型がどのように共存しているかを調べることは非常に興味深いことである。
今回の実習の目的は以下の2点である。
ベニシダ類の有性生殖型と無配生殖型が屋久島の中でどのように生育しているのか、生育している海抜高度や微環境が異なることで共存しているのかどうかを調べる。
胞子嚢あたりの胞子数を計数することで薄嚢シダ類の生殖様式を識別する方法を学ぶ。また、マッピング法による調査を実際に体験し、野外で植物の生育環境を調べる方法も学ぶ。
(マッピング法とは、ある植物が生育している位置を座標軸にプロットとすることで、その植物の分布と生育している環境を調べることである。)
今回材料としたベニシダ類とは、オシダ属(Dryopteris, 基本的に丸い胞子嚢群をもち、それが円腎形の包膜でおおわれ、羽片や葉脈は外先の分岐をすることで特徴づけられる属)の中で葉に袋状の鱗片があり、葉質が紙質の(うすい)シダ植物である。ベニシダ類には種の識別が難しいものが多いが、群としては良くまとまった比較的認識しやすい1群のシダ植物である。日本では、およそ20種類が分類学上の「種」や「変種」として扱われているが、そのほとんどが無配生殖型で、有性生殖型は2倍体であるホコザキベニシダなど、3種類しか確認されていない。無配生殖型は、調べられた限りでは大部分が3倍体である。無配生殖型の一部は、形態的に他の種類の間の中間型を示しており、交雑が関係する複雑な種分化が起こってきた可能性がある。
今回はまず、屋久島に生育するベニシダ類の有性生殖型と無配生殖型で生育する標高(海抜高度)が異なっているかどうかを調べるために、モッチョム岳の標高約200mの登山道入り口から標高約750mの万代杉付近までの間に生育しているベニシダ類、及び安房林道沿いの標高約400mから標高約1200mまでに生育しているベニシダ類について、適当な間隔で個体を選び、その個体の生育地点の標高およびまわりの環境を記録し、通し番号を記したビニールテープをその株に貼付け、胞子のついた葉を1枚採集して、採集した葉にもビニールテープで同じ番号をつけた。
採集したそれぞれの葉から胞子嚢を1つ採取し、その中に含まれている胞子の数を数えることにより、その個体の生殖様式(有性生殖であるか無配生殖であるか)を判別した。先に「はじめに」のところで無配生殖を説明した際にも述べたように、薄嚢シダ類の有性生殖型では、胞子嚢あたりの胞子の数が64個であるのに対して、無配生殖型では32個となる(図3)。

図3 上-有性生殖型の胞子(n=64)
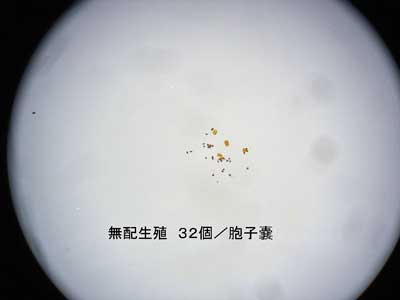
図3 下-無配生殖型の胞子(n=32)
したがって胞子嚢あたりの胞子の数を数えれば、その個体の生殖様式を容易に推定することができるのである。
胞子嚢あたりの胞子を数える具体的な方法としては、まず胞子嚢群から柄付き針で破裂していない胞子嚢をひとつ取り出し、スライドグラスの上にのせ、カバーガラスをかぶせる。そしてカバーガラスの上から指で軽く叩いてやると、胞子嚢が壊れ、中の胞子が飛び出し、胞子数を数えることが可能となる。誤判別を減らす為に一個体あたり最低でも2個の胞子嚢について胞子を数えた。胞子の観察には、携帯用実体顕微鏡(Nikon社、ファーブル)を用いた。
モッチョム岳において無配生殖型と有性生殖型が同所的に混生して生育していた地点については、番号札を頼りに、その地点に戻り、その近辺に生育している全成熟個体(胞子を付けている個体)について個体別マッピング調査を行なった。マッピング調査の具体的な方法は、歩道沿いに50mの巻き尺をはって、これをX軸とし、それと垂直方向をY軸とした。Y軸方向は、2mの金尺(コンベックス)を使い、50mの巻き尺の山側を+、逆の谷側を-として、巻き尺の両側の合計4mの範囲に生えている全てのベニシダ類の成熟個体の位置をXYの座標として記録した。そして先の調査同様、記録した全ての株について通し番号を記したビニールテープを貼付け、それぞれの個体から胞子のついた葉を1枚採集して、採集した葉にもビニールテープで同じ番号をつけた(図4)。

図4 モッチョム岳における調査風景
採集した葉については持ち帰って宿舎で胞子嚢あたりの胞子数を数え、各個体の生殖様式を推定した。さらに異なる生殖型間の生育環境にどのような違いがあるかを詳しく調べるために、生殖様式の推定が終わった段階で、再度同じ地点に戻り、株に付けた番号を頼りにして推定した生殖様式で色分けしたテープを各個体に貼ることにより、有性生殖型と無配生殖型がどのように生えているかが一目でわかるようにした。その上で、生育環境に比較をおこなった。
胞子嚢あたりの胞子数を数えるために採集した葉については、すべてさく葉(押し葉)標本とし、そのさく葉標本を用いて様々な形態形質の比較を行った。
1.生殖様式と生育する標高の関係
モッチョム岳、安房林道、共に有性生殖型は比較的低い標高に多く成育しており、モッチョム岳、安房林道のいずれにおいても標高約400m~600mの間に最も多く生育していた(図5)。
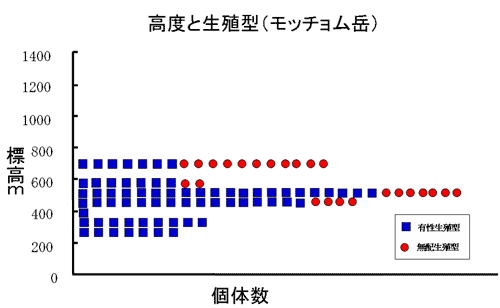
図5 生殖様式と高度の関係-モッチョム岳
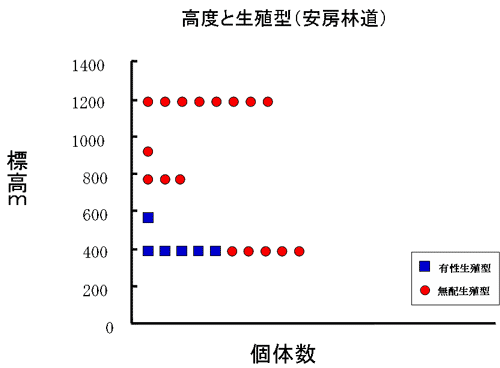
図5 生殖様式と高度の関係-安房林道
安房林道の高度600m以上では、有性生殖型はみられなかった。逆に無配生殖型は、モッチョム岳、安房林道共に高度が上がるにつれて観察される個体数が多くなっていった。有性生殖型と無配生殖型の混生集団はモッチョム岳では標高540m付近で、安房林道では標高400m付近でみられた。
2.有性生殖型と無配生殖型の混生集団のマッピング調査
ベニシダ類の有性生殖型と無配生殖型が混生しており、それぞれの個体数も多かったモッチョム岳の標高540m付近の集団について全成熟個体の個体別マッピング調査を行った。林道沿いに縦50m 横4mのコデラートを張り、コデラート内のベニシダ類全成熟個体についてその生殖様式を推定して、現地で生殖型ごとに異なる色のビニールテープで色分けした。その結果、この場所では有性生殖型と無配生殖型は完全に混生しているように見え、特に生殖型間で生育環境の差異は観察されなかった(図6、7)。
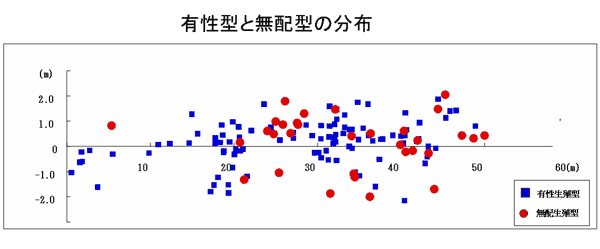
図6 マッピング調査地内における有性生殖型と無配生殖型の分布


図7 有性生殖型と無配生殖型の混生の様子
3.各生殖様式にみられた形態形質 しかし、これらの形態型の間には、様々な中間的形態を有する個体が存在し、どの型に属するか解らない個体も観察された。さらに少数ながら、有性生殖を行う個体でも無配生殖型に多くみられるような形態形質を有する個体や、無配生殖を行う個体でも有性生殖に多くみられるような形態形質を有する個体も観察された。
4.形態型と生育標高の関係 図15 各形態型と高度の関係-モッチョム岳 図15 各形態型と高度の関係-安房林道 5.マッピング調査地における各形態型の分布 図16 マッピング調査地内の各形態型の分布 ベニシダ類の有性生殖型と無配生殖型は、屋久島において海抜高度によるすみわけを行なっている可能性が示唆された。モッチョム岳、安房林道のいずれにおいても、有性生殖型が主として標高約400m?600mの間に生育していたのに対して、それ以上の標高では無配生殖型の方が徐々に多くなっていくという傾向が今回の調査結果において見られたからである。一方で無配生殖と有性生殖が同所的に混生して生育する集団も存在し、その集団ではかなり詳細に生育環境の比較を行ったにもかかわらず、無配生殖型と有性生殖型に生育環境の差異は観察されなかった。というよりも、今回調査したコデラート内において、どの地点においても有性生殖型と無配生殖型は、まさに隣り合うように生えており、生態的分化があるとは到底思えない状況であった。ただし、大きく見れば有性生殖型が好んで生える標高と無配生殖型が好んで生える標高に違いがあることが示唆される結果もえられているわけで、それぞれの生殖型にとって好適な環境(温度など)は異なっていることが予想される。したがって、今回コデラート調査したような混生集団が維持される(「はじめに」のところで述べたように、生態的分化のない有性生殖型と無配生殖型が共存すると、増殖力にまさる無配生殖型があっという間に多数派を占め、有性生殖型を排除してしまうと考えられる)理由を説明するひとつの仮説としては、モッチョム岳の540m付近の環境では、無配生殖型の最も好適な生育環境ではないため、有性生殖型が無配生殖型と同所的に生育できている可能性が考えられる。実際、今回のコデラート中では、有性生殖型の方が多数派を占めていた。無配生殖型の方が多数派を占めるような集団で2つの生殖型がどのようになっているかは興味深い。 京都大学野生動物研究センター>屋久島フィールドワーク講座
>第7回・2005年の活動-シダ班-報告書 このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹
さく葉標本を用いた無配生殖型と有性生殖型の形態形質の比較の結果、それぞれの生殖型にみられた典型的な形態型の間には葉の形に差異がみられた。また無配生殖型の中には大きな形態変異がみられ、複数の形態型が認められた。
有性生殖型は葉の頂上部が急に細くなり、頂羽片状となるもの(いわゆるホコザキベニシダ型)が大多数を占めた(図9)。この型では、葉の中軸と羽片の軸は斜めに交わり、最下羽片の下向き第一小羽片は第二小羽片よりも小さくて切れ込みは浅く、胞子嚢群は羽片の基部側に偏る傾向があり、小羽片の軸と縁の中間か、やや軸寄りにつく。若い葉は赤みを帯びるが、成熟すると葉身も葉軸も緑色になる。
無配生殖型には、以下のような4つの形態型が見られた。葉軸が紫色になり、羽片の軸は中軸に対して直角に近く、胞子嚢群は小羽片の縁寄りにつく型(ムラサキベニシダ型、図10、11)。葉軸は緑色で、羽片の軸と中軸がほぼ直交し、羽片は鎌状に斜上する型(ヌカイタチシダモドキ型、図12)。胞子嚢群は羽片の先端側に偏る傾向があり、最下羽片の下向き第一小羽片が第二小羽片長くてより極端に羽状に分裂し、葉身が五角形のようになる型(タカサゴシダ型、図13)。ホコザキベニシダ型とタカサゴシダ型の中間的で、最下羽片の下向き第一小羽片は第二小羽片よりやや小さいか、またはやや大きくて深く切れ込む型(トウゴクシダ型、図14)。



図9 ホコザキベニシダ型 図10 ムラサキベニシダ型(表) 図11 ムラサキベニシダ型(裏)



図12 ヌカイタチシダ型 図13 タカサゴシダ型 図14 トウゴクシダ型
有性生殖型にみられたホコザキベニシダ型はモッチョム岳、安房林道共に標高400?600mまでの間に最も多く観察された。無配生殖型にみられた形態の型のうちタカサゴシダ型はモッチョム岳でも安房林道でも高度の高い地点にのみ生育していた。トウゴクシダ型は、モッチョム岳では標高500m付近で最も多く生育しており、安房林道においては標高800m以上に多く生育していた。ムラサキベニシダ型は、モッチョム岳では標高500mと750mにおいて、また安房林道では標高400m付近で多く生育していた。ヌカイタチシダモドキ型は、モッチョム岳の標高500mから750mにかけて生育していた(図15)。


今回マッピング調査したモッチョム岳の標高540m付近のベニシダ類集団において、無配生殖型の各形態型(個体数の多かったムラサキベニシダ型、ヌカイタチシダモドキ型)も完全に混生しており、無配生殖型の各形態型間でも生育環境の違いは特に見出されなかった(図16)。
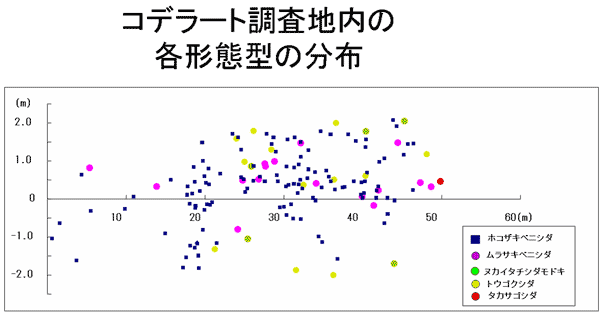
考察
形態の比較・解析の結果、有性生殖型は葉の形にまとまりがみられた。少なくとも、今回調べた有性生殖型の大部分は、ホコザキベニシダと呼ばれる形態型であり、これらは1つの実体と考えられる。
一方、無配生殖型には複数の形態型が観察され、起源の異なる複数の実体が含まれている可能性が強く示唆された。無配生殖型シダ類の起源に関する過去の研究では、オシダ属、チャセンシダ属、イノモトソウ属などで、無配生殖型が有性生殖型の祖先から進化してきたことが明らかにされている。その中には、単一の有性生殖型から起源したと考えられるものも、形態的、系統的に異なる2つの有性生殖型、あるいは有性生殖型と無配生殖型の間の交雑に由来すると考えられるものもある(無配生殖型が、まれに父親あるいは母親として、配偶子を形成して他個体と交配し、有性的に次世代を残す現象が報告されている)。無配生殖型のうち、タカサゴシダ型とヌカイタチシダモドキ型は、形態的にホコザキベニシダ型とは多くの点で異なっており、また互いに異なってもいる。このことは、それらの無配生殖型がホコザキベニシダ型とは形態的に(おそらく生態的にも)異なる少なくとも2つの異なる有性生殖型から、それぞれ起源した可能性を示唆している。タカサゴシダ型に関しては、台湾で有性生殖型が観察されており、これが屋久島産無配生殖型の祖先の候補と考えられる。ヌカイタチシダモドキ型についても、未発見ないし絶滅した有性生殖型の祖先があると推測される。ムラサキベニシダ型は、葉の中軸と羽片の交わりが直角に近い点ではヌカイタチシダモドキ型と似ているが、葉の先が細くなってホコザキベニシダ型に似るものもある。また、トウゴクシダ型は最下羽片の下向き第一小羽片がやや大きく、小羽片の切れ込みが深い点でタカサゴシダ型に似ているが、やはり葉の先が細くなるものがある。ムラサキベニシダ型とトウゴクシダ型は有性生殖型(ホコザキベニシダ型)と無配生殖型(ヌカイタチシダモドキ型とタカサゴシダ型)との中間的な形態と見ることもできる。それらの起源には、なんらかの交雑現象によって、ホコザキベニシダ型が関わっている可能性が示唆される。
典型的な形態型の間に、様々な中間型が存在する現象については、次のような仮説が考えられる。(1)それぞれの形態型に変異の幅があって、その中のあるものが、たまたま中間的な形をしているに過ぎない。(2)無配生殖型と有性生殖型、あるいは形態の異なる無配生殖型の間のなんらかの遺伝子交流(交雑?)の結果、中間的な形質をもつ個体が造られている。これらの仮説を検証するには生殖様式の調査の他に細胞学的、および遺伝学的解析(染色体の観察、酵素多型、DNAなどの分子マーカーを用いた研究)が必須であり、今後、これらの研究がおこなわれることによって屋久島のベニシダ類の多様性の由来について、より理解が深まることが期待される。