図1-1 2007年8月23日GPSデータによる遊動地図
各群れの線は5分ごとの観察者の位置をつないだもの。受信状態の悪い時は精度が低く、実際の移動とはことなる飛び離れた位置を示すことがある。
図1-2 2007年8月24日GPSデータによる遊動地図


図1-3 2007年8月25日GPSデータによる遊動地図
図1-4 2007年8月23日GPSデータによる遊動地図 この日は3群を同時追跡した。われわれがE群とNAⅠ群を、京大霊長類研究所の杉浦秀樹博士がNA2群を追跡した。
| 京都大学野生動物研究センター>屋久島フィールドワーク講座>第9回・2007年の活動-サル班-報告書 |
| 概要 | 人と自然班 | サル班 | シカ班 | 植物班 | 公開講座 | スタッフ |
| 報告書 | 感想文 |

講師: 鈴木 滋、西川 真理
参加者: 小路田俊子・佐伯美幸・綿引和巳・林恵理子
ヤクシマザルは、ニホンザルの亜種である。特徴としては以下のようなものがあげられる。
今回の調査では、我々は群れ間の関係について注目することにした。ヤクシマザルは隣り合った群れと行動域が重複しているため、群れ同士が出会う頻度が高い。そこで、「隣り合った群れ同士が出会った時あるいは接近したときに、サルの行動にどのような変化が現れるのか」を2群同時追跡することによって調査した。
屋久島のサルの群れ同士の関係の基本は、敵対的で、強い群れと弱い群れがはっきりとしており、群れ同士が出会うと対峙したり、弱い方が強い群れに追われたりすることが知られている。そこで、群れは、隣接群と出会ったり、出会いそう場所では、いつもとは違う行動をとるのではないかと考えた。
「隣り合った群れ同士が接近した時に、サルの行動にどのような変化が現れるのか」についての具体的な行動の予想は次のようになった。
半山地域において隣接する、ヤクシマザル(Macaca fuscata yakui)のE群とNA2群、NA1群を調査対象とした
西部林道半山地区(南稜~北稜)
調査区域である半山に生息している植物の名前と特徴を学びつつ、散策。サルの食べ物やサルの生活環境を体感した。
データを取る練習をした後、実際にデータを取った。E群はギンを10時~15時まで、NA2群はナガラメを10時55分~15時まで追跡した。
E群はギンを10時05分~16時30分まで、NA2群はサンゴジュを10時05分~16時30分まで追跡した。
E群はミナミを10時~16時30分まで、NA2群はノリコを10時~16時30分まで追跡した。
E群はカエルを9時~11時30分まで、NA1群はホタルを9時~11時30分まで追跡した。(この日はE群の近くにNA1群がいたので、調査対象をNA2からNA1に変更した。)
サルの観察を2つのグループにわかれて行った。観察対象の2つの群れに、それぞれ学生2名と講師かチューターが1名つき、群れのオトナメス1個体を追跡個体に決めて、個体追跡法を用いて活動データを収集した(表1)。
表1 追跡群と個体追跡対象個体と追跡時間
| 群れ | 個体名 | 日付 | (分) |
| E | ギン | 2007.8.23 | 305 |
| ギン | 2007.8.24 | 390 | |
| ミナミ | 2007.8.25 | 395 | |
| カエル | 2007.8.26 | 155 | |
| NA2 | ナガラメ | 2007.8.23 | 250 |
| サンゴジュ | 2007.8.24 | 395 | |
| ノリコ | 2007.8.25 | 460 | |
| NA1 | ホタル | 2007.8.26 | 170 |
| 総計 | 2520 |
それぞれのグループの学生2人のうちのひとりは、活動を採食、移動、休息、毛づくろい、その他(喧嘩・交尾・遊びなど)に分け、5分毎に記録した。また、採食の植物や動物の種類と採食部位も記録し、樹木で採食した場合はその樹木のナンバーテープの番号を記録した。
グループの学生の2人目は、GPSを携行し、位置データを5分毎に自動記録するとともに、DOP値と受信状態(未受信/2D/3D)を記録した(図1)。なお、群れ間の距離の分析に使用したデータの信頼度は3Dで,信頼性の高いもののみを分析の対象とした。
また、5分毎に鳴るアラームから1分以内に、追跡個体と群れの他個体のそれぞれにおいて発声があるかどうかと、追跡個体のオトナメスの近くにオトナオスがいるかいないか、いる場合どのくらいの距離でいるのかを記録した。
私たちが4日間で追跡したのは1個体だが、その個体で個体の属する群れを代表させて分析した。
個体追跡をした4日間に集めたデータは、GPSの位置データ、ヤクシマザルの5分毎の活動、採食品目、近接オス、発声頻度である。これらを使い、群れ間の距離と、採食品目、活動内容、近接オスの頻度、音声頻度、応答期待値、エンカウンター直後とそれ以外の音声頻度の分析を行った。
近接オス頻度と、音声の頻度については、個体の凝集性と、音声のやりとりの積極性の指標とするために、以下のようにポイントを設定した。
もっとも近いオトナのオスが、
5分毎の観察のはじめ1分間に
また、群れ間の距離との関係を分析するとき、毛づくろいと休息時のデータはのぞいて、近接オスポイントを計算した。これは、毛づくろいと休息(毛づくろいの合間であることが多かった)で、ほとんどの場合群れの個体が1カ所に集まっているため、群れ間の距離によってオスの近接の程度が変わることを分析するのには、適さないデータだと考えるからである。
|
|
図1-1 2007年8月23日GPSデータによる遊動地図
各群れの線は5分ごとの観察者の位置をつないだもの。受信状態の悪い時は精度が低く、実際の移動とはことなる飛び離れた位置を示すことがある。 |
図1-2 2007年8月24日GPSデータによる遊動地図 |
|
|
図1-3 2007年8月25日GPSデータによる遊動地図 |
図1-4 2007年8月23日GPSデータによる遊動地図 この日は3群を同時追跡した。われわれがE群とNAⅠ群を、京大霊長類研究所の杉浦秀樹博士がNA2群を追跡した。 |
GPSのデータをもとに、8月23,24,25,26日の追跡個体間の距離、すなわち群れ間の距離について分析した(図2)。群れの組合せごとにわけず、すべてプールして分析した。
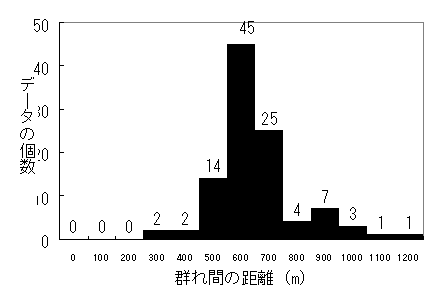
図2 群れ間の距離の分布 数字は5分ごとに1回の記録サンプル数
隣接群とは平均603mの距離をとっていた。最大1135m、最小294mであった。
この間に、群れ同士がエンカウンターしたと思われるのは、以下の最終日の観察当初のみである。それ以外の期間は、他群の音声や個体を明瞭に観察することはなかった。
最終日の3群同時追跡のとき、E群とNA1群は、おそらく出会った後に正反対の方向に離れるように移動をした。また、どちらの群れも3つ目の群れであるNA2群に気づいていそうな距離におり、あとから遊動状況を照らし合わせると、NA1群は、NA2群も避けるように長距離直線的な移動をしたようにみえた。この日の群れ間の距離をとりだして、群れの組合せごとにわけてみたのが図3である。もっとも接近していたNA1群とNA2群の間は、300m以下では、隣接群の音声が確認できるが、群れ間が400mの距離でNA1群の位置は安定していた。またEとNA2群間では、前日までと同様600m程度の距離をあけて安定していた。E群とNA1群の距離は900mで大きいが、位置関係としては、2つの群れの間にはNA2群がいる状態が長かったので、隣接していたとはいえない。
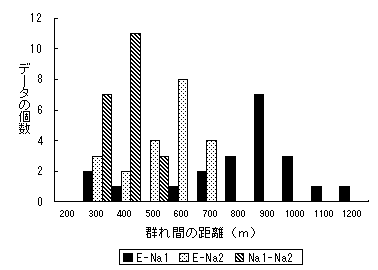
図3 3群同時追跡した日の群れ間の距離 遊動状況
図1?3を参照のこと
これらの結果をふまえて、群れ間の距離の近いか遠いかで行動がどのように変化するのかを分析するために、以下のように群れ間の距離のクラスわけをした。
| 群れ間距離 | 距離段階 | データ数 |
| 0~499m: | 近距離 | 18 |
| 500~699m: | 標準距離 | 70 |
| 700m~ : | 遠距離 | 16 |
群れ同士が接近している時とそうでない時とを区分するために、分析によっては標準と遠距離をあわせて1つの段階「近距離外」とした。
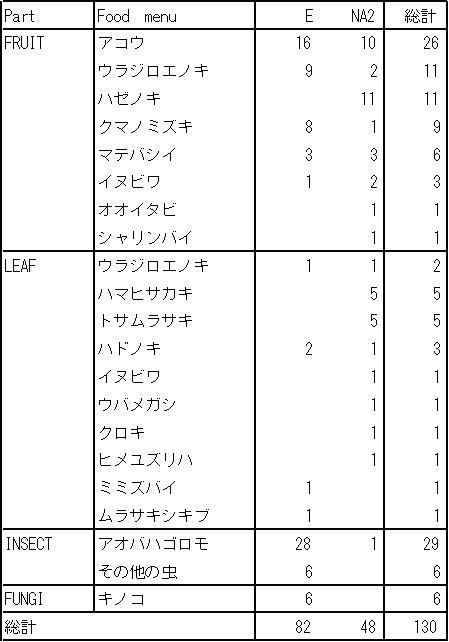
NA2群とE群について、採食品目ごとのデータ数を8月23,24,25日について比較した。(26日は片方のグループがNA1群を追ったため比較から外した。)
4日間で食べた食物はおよそ25種類であった。3日間ともE群の方が食べている時間が長かった。
アコウの果実、ウラジロエノキの果実、は両群ともよく食べているが、他の品目についてはあまり共通性は見られなかった。E群はアオバハゴロモ、アコウの果実が多く、NA2群はアコウの果実、ハゼノキの果実が多かった。
採食対象が、群れ間の距離によってことなるかどうかは、データ数が少ないために検討できなかった。
表1 追跡群と個体追跡対象個体と追跡時間
群れ間の距離が近いと、群れの活動パタンには変化があるのだろうか。このことを検討するために、群れ同士が近いときと、そうでない時の2段階にわけて、活動パタンを比較した。ここでは、行動を採食、毛づくろい、移動、休息の4つのカテゴリーにわけ、それぞれに対し、距離段階ごとのデータ数の割合を示した(図4)。なお、分析には8月23,24,25日のデータを使用した。ここで、群れ間の距離が「近距離外」とは標準距離と遠距離のデータを足したものである。
どちらの群れも、群れ間の距離にかかわらず、4つの活動の中で毛づくろいか休息がもっとも多くみられた。
とはいえ、群れの同士の距離によって活動パタンの割合には違いがみられた。どちらの群れでも、採食は、遠距離よりも近距離でよく行われたが、毛づくろいは近距離よりも遠距離でよく行われた。移動と休息には群れ間に共通の傾向は見られなかった。
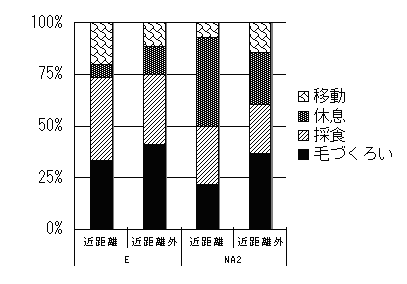
図4 群れ間の距離と活動パタン割合
群れ間の距離が近い場合と、近くない場合とで、追跡個体のメスとオスの近接度を比較した。ここでは、毛づくろい、休息時のデータは除いた。8/23,24,25,26のデータを使用。
E群、NA2群とも群れ間距離が小さい場合のほうが、メスとオスが近接していることが多かった。
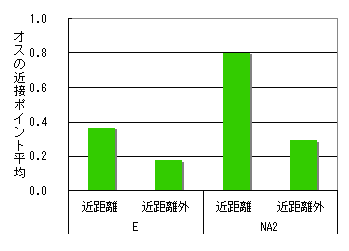
図5 群れ間の距離とオスとの近接の程度 (移動・採食時のみ)
両群とも群れ間距離が小さい時のほうが、声をよく出していた。追跡個体が移動採食中の場合(図6)でも、休息毛づくろい中(図7)でも同じ傾向がみられた。
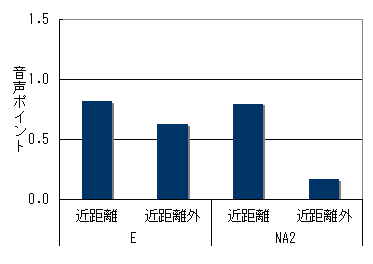
図6 群れ間の距離と音声の頻度(移動・採食時)
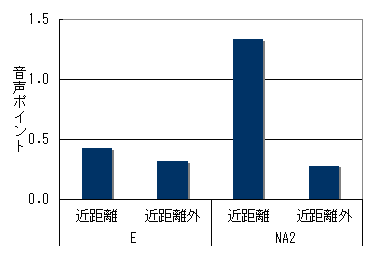
図7 群れ間の距離と音声の頻度(休息・毛づくろい時)
また、群れ間距離が近いときと近くないときで、発声頻度と群れ内の仲間からの応答率を比較したところ、発声の観察率のみならず、応答があった割合も群れ間距離が近い時のほうが高く、より鳴き交していたことがわかった(表3)。
表3 群れ間の距離と発声割合と応答割合
| 近い | 近くない | 全体 | |
| 発声率 | 51% | 26% | 31% |
| 応答率 | 61% | 44% | 49% |
発声率=(音声ポイント1,2のデータ数)÷(音声ポイント0,1,2のデータ数)×100
応答率=(音声ポイント2のデータ数)÷(音声ポイント1,2のデータ数)×100
8月26日のエンカウンター直後(9:00~9:30)とそれ以後(9:35~11:30)で発声頻度を音声ポイントをつかって比較した(表)。どちらの群れでも、エンカウンター直後はそれ以後に比べ発声頻度が下がっていた。また、エンカウンター直後、それ以後ともE群の方がよく発声していた。
表3 群れのエンカウンターと発声頻度
| 群れ | 直後 | それ以降 |
| E | 0.29 | 0.60 |
| NA1 | 0.14 | 0.40 |
| 両方 | 0.21 | 0.46 |
エンカウンター直後とそれ以後で、オスの近接ポイントの平均値を比較した(表4)。E群ではエンカウンター直後のオスの接近は見られなかった。 NA1群ではエンカウンター直後にオスが近くにいることが多かった。
表4 群れのエンカウンターとオスとの近接
| 群れ | 直後 | それ以降 |
| E | 0.00 | 1.87 |
| NA1 | 2.14 | 2.05 |
| 両方 | 1.07 | 1.95 |
隣り合った群れが出会うことを「エンカウンター」と呼び、そこではお互いの群れが豊富な食物資源を供給する遊動域を巡って、個体間の優劣と同じように、攻撃的な交渉がおこり、群れ間にも優劣があるとされている(丸橋,1991)。今回の調査では、残念ながらエンカウンターしている現場を直接観察することはできなかったが、エンカウンターをしたのではないかと思われるほどE群とNA1群が接近している時の同時追跡をすることができた。さらに、隣接2群間の距離はおおよそ600メートル前後であり、その距離が近くなれば、発声頻度やオスとの接近に影響があることが示唆される結果をえた。
サルは、およそ群れ間距離が500m程度から隣接群の位置を認識できるのではないかと思われる。500m程度離れていると、木が生い茂った森の中で隣接群を目で発見することは困難であると考えられる。一方、500m程度であればサルの発する音声は届くのかもしれない。実際、遠くからサルの音声が聞えた時に、トランシーバーで同時追跡している別の班にその班が追跡している班が発声したものか確認したところ、500m程度であった事があったので、少なくとも比較的大きな発声であれば隣接群も聞き取ることができるのかもしれない。大井他(2003)は、屋久島の同地域でサルの音声の探知確率を調べているが、ラウドコールと悲鳴は400mぐらいまでは探知確率が落ちないが、500m前後で急減し、最大可聴範囲が580mであったことを報告しており、今回の推測を裏づけている。
隣接群との距離が500mより近い場合よりもそれ以上である状態の方が明らかに多かったが、近距離とそうでないときでは発声頻度やオスとの近接の程度が異なっていた。しかし、サルが隣接群との位置関係の認識に聴覚情報を頼っているかどうかは今回の調査では明らかにできない。というのも、経験的にエンカウンターしそうな場所を知っているから行動に変化があっただけかもしれないからである。
次に、どうして隣接群との距離が小さければ発声頻度が高くなるのだろうか。サルが音声を発する目的のひとつは、群れ内の各個体の位置情報を共有することと考えられる。ヤクシマザルは、他の群れが接近すると、群れ内でのコミュニケーションを活発にすることによって群れの結束を強くし、他の群れに対抗しようとしているのではないだろうか。
隣接群が近い場合ほどオスとメスが近接していたのはどうしてだろうか。隣接群とエンカウンターした時に強いオスがいるほうがほかの群れよりも優位になり、より食物資源がゆたかな遊動域を確保できるだろう。他群との接近時にはオスにかぎらず、群れ内の個体間の距離が近くなっている可能性があるが、今後の課題である。
群れ間には採食をめぐる競合があると考えられるが、群れ同士の接近時には、サルは採食により積極的になるのかもしれない。群れ間の距離が近いと、どちらの群れでも採食割合が高く、また、毛づくろいの割合が低かった。これまでにも、群れ間の出会いは、しばしば大きな採食パッチをめぐって起こりやすく、優位な群れが劣位な群れを立ち退かせて、採食パッチを占有することが知られている(丸橋,1991)。
最後に、エンカウンターしたのではないかと思われる8月26日のデータについて考察する。このときの群れは3群が同時に接近しており、観察終了後に移動のトレースをつきあわせた結果、NA1群がNA2群とE群を避けて移動し、E群がNA2群を避けて移動したと考えられた。つまり群れ間にはNA2群>NA1群>E群の順序で優劣があったように思われる。強い群れをさけたNA1群とE群は、エンカウンターの直後は発声するのを控えたのではないだろうか。また、E群ではオスが近くにはきていなかったが、NA1群ではエンカウンター直後にオスが近くにきていた。これは弱い群れではよりオスの近接がメスにとって重要であるからかもしれない。
以上をまとめると、西部林道のヤクシマザルは、隣接群との距離が500m未満になると群れの存在を何らかの形で把握し、発声頻度を増加させることにより群れ内のコミュニケーションを深め、群れ内にオスとメスが近接することにより隣接群とのエンカウンターに備えていると考えられる。
今回の我々の研究結果では、サルは音声によって群れ内のみならず他の群れに存在をアピールすることももたらしている可能性が示唆された。群れ同士が近くにいるときに、発声頻度が高まっていたこと、さらに、丸橋珠樹(1991)によると、採食中の場合、ヤクシマザルは発声によりお互いの位置を確認することで他個体と接近して採食パッチ内で出会うことを避けるために使っているとされることから、音声は、群れ間の距離を調整するためにも使われているのかもしれない。
今回の結果は、データ数が少ないために結論的なことはいえないが、個体の音声や採食行動が、他群存在によって影響をうけている可能性をみいだすことができた。今後、群れの同時追跡を続けることによって、群れの分散度や他個体と採食競合などに、隣接群からの影響があるのかどうかなどを検討できると思われる。
音声を手がかりとしたニホンザル野生群の位置探索の正確さについて 霊長類研究,19: 193?201.
「群れの社会変動論?屋久島のニホンザル社会生態?」『サルの文化誌』(西田・伊沢・加納編)平凡社,東京.pp. 109?127.
今回の調査にあたっては、屋久島森林管理署と鹿児島県より入林許可等の便宜をいただいた。また、Topher Dagg 氏から、NA2群の個体情報や遊動域について詳細な情報を提供していただいたうえに、ご自身の研究について、スライドをつかったわかりやすい紹介をしていただいた。また、霊長類研究所の杉浦秀樹博士には、サルの個体識別情報をはじめ、さまざまな有益な情報を提供していただき、また、研究内容についてもフィールドでご教示いただいた。記して感謝したい。
京都大学野生動物研究センター>屋久島フィールドワーク講座 >第9回・2007年の活動-サル班-報告書
このページの問い合わせ先:京都大学野生動物研究センター 杉浦秀樹