Empathy and Fairness: Psychological Mechanisms for Eliciting and Maintaining Prosociality and Cooperation in Primates
Shinya Yamamoto, Ayaka Takimoto
DOI: 10.1007/s11211-012-0160-0Abstract
In the past, prosociality has been considered a hallmark of humans; however, recently, accumulating data have empirically revealed that non-human animals also show prosocial behavior. In situations in which animals cannot predict return benefits, prosocial behavior is probably driven by other-regarding motivation. A sense of fairness and empathy continue to draw attention as the most plausible candidates for the psychological mechanisms underlying such prosocial behavior. In this article, we first introduce comparative studies on prosocial behavior in non-human primates and discuss similarities and differences between humans and non-human primates. Then, we discuss the role of a sense of fairness and empathy. In this paper, we hypothesize that empathy may promote prosocial behavior, whereas a sense of fairness may play a role as a stabilizer, but not as a promoter of prosocial behavior in non-human animals. We further hypothesize that prosocial behavior motivated by sympathetic concerns can survive only with a sense of fairness, the inhibitory system for unnecessarily excessive expression of prosocial behavior. Without a sense of fairness, empathic animals might be exploited by free-riders, which might lead to the extinction of cooperation. Therefore, the interplay of a sense of fairness and empathy are both important to maintaining prosocial behavior and cooperation. This hypothesis seems to be supported by comparative studies with non-human primates and also by neural studies with humans.
Introduction
In this pair of special issues, “Justice in animals,” a sense of fairness that appears in the form of a behavioral response of inequity is the central discussion topic. Why has fairness drawn so much attention? It may be because fairness is closely related to human cooperation and prosociality. Although, in classic economic theories people are assumed to be exclusively motivated by their own self-interest, researchers have noticed that in actuality, humans are also strongly motivated by other-regarding preferences, or interest in others’ outcomes. From this perspective, fairness has become the focus of modern economic theories explaining human prosociality (Fehr & Schmidt, 1999). If this is the case in humans, it may also be true in other species that have recently been shown to demonstrate prosocial behavior. Thus, non-human animals as well as humans have become a target for investigation of a sense of fairness, and researchers have devoted efforts to finding the evolutionary link among prosociality, cooperation, and fairness (Brosnan, 2011).
We place prosociality at the center of the discussion in this article. Prosocial behavior refers to “voluntary actions that are intended to help or benefit another individual or group of individuals” (Eisenberg & Mussen, 1989). This definition refers to the consequences of a donor’s actions rather than the motivations behind those actions. These behaviors include a broad range of activities, such as: sharing, comforting, rescuing, and helping. If there is a cost to the actor, then the behavior can be labeled as altruistic behavior; therefore, prosocial behavior includes altruistic behavior (Yamamoto & Tanaka, 2009a).
Previously, the evolution of prosocial behavior was mainly explained from an ultimate perspective, i.e., the cost–benefit balance of the behavioral consequence. Recently, however, prosocial behavior has been analyzed from the proximate viewpoint of psychological mechanisms (Brosnan, 2011; de Waal, 2008; Preston & de Waal, 2002). From an ultimate perspective, it is argued that prosocial behavior might subsequently bring some benefits to the actor, such as through reciprocal interaction (Trivers, 1971) and helping kin (Hamilton, 1964). However, the underlying motivations are not necessarily based on self-interest. When an individual cannot predict any return benefit, the prosocial behavior must be driven by other-regarding motivations (de Waal, 2008; de Waal & Suchak, 2010). Empathy, the ability to share the feelings and emotions of others, as well as a sense of fairness, are considered to be the most plausible underlying psychological mechanisms of this other-regarding behavior.
Here, we also define another closely related term, “cooperation.” This word has acquired two different meanings, which has caused some confusion among researchers in different fields. In some disciplines, such as primatology and ethology, this term is often used as “collective action”: the behavior of two or more individuals acting together to achieve a common goal, typically in hunting context (Busse, 1978; Boesch & Boesch, 1989) and bar- or rope-pulling cooperative tasks (Chalmeau, 1994; Crawford, 1937; Hirata & Fuwa, 2007). With this definition, cooperation can be achieved by only self-regarding motives. In other research, “cooperation” is used in the analysis of individual behavior: an individual acts cooperatively only if it benefits the long-term fitness (in terms of survival and reproduction) of both itself and the recipient of its actions, even at the expense of its own short-term fitness (Hamilton, 1964; Krebs and Davies, 1987; Maynard-Smith, 1982). Melis and Semmann (2010) adopted this term in broader sense along with this line: behaviors which provide a benefit to a recipient or are beneficial to both the actor and the recipient. In this usage, by definition, cooperation is compatible with prosocial behavior. In this article, we follow this latter definition, and use “cooperation” when discussing reciprocal cooperation (i.e., reciprocal prosocial or altruistic behavior). When we refer to cooperation based on the former meaning, we use “collective action.”
Previously, prosocial behavior, a sense of fairness, and empathy were considered to be some of the hallmarks of humans. However, recently there has been accumulating evidence for the existence of these traits in non-human animals as well. In this article, we first introduce empirical studies suggesting similarities and differences between humans and non-human animals in prosocial behavior and mechanisms related to fairness and empathy. Following this, we discuss how each of the two psychological traits, a sense of fairness and empathy, impact prosocial behavior; finally, we also discuss the possible link between fairness and empathy.
Comparison of Studies on Prosociality with Human and Non-human Animals
Many non-human species in different taxa have recently been shown to demonstrate prosocial behavior in the wild and captivity (de Waal, 2008; Dugatkin, 1997: Kappeler & van Schaik, 2006; see also Price & Brosnan, 2012, and Bräuer & Hanus, 2012). Recently, two experimental paradigms have been developed to empirically test non-human animals’ prosociality: one is a prosocial choice paradigm and the other is a targeted helping (or instrumental helping) paradigm.
In the prosocial choice paradigm, an animal is presented with opportunities to provide benefits to others at little or no cost to themselves (Fig. 1a). The animal can choose one of two options: one produces rewards to him/herself and a conspecific partner (prosocial choice), and the other benefits only him/herself (selfish choice). Whether the animal chooses the prosocial or selfish choice, he or she can obtain the same amount of reward; therefore, giving a benefit to the partner by choosing the prosocial option is not costly, although it may require a little labor (e.g., pulling a bar connected to the food tray). This prosocial choice test is now one of the most prevalent test paradigms applied to various species and has revealed considerable species differences (Table 1). A prosocial tendency, i.e., the tendency for an actor to choose the prosocial choice more frequently when the partner is present than absent, has been observed in New World monkeys (capuchin monkeys: de Waal, Leimgruber, & Greenberg, 2008; Lakshminarayanan & Santos, 2008; Takimoto & Fujita, 2011; Takimoto, Kuroshima, & Fujita, 2010, common marmosets: Burkart, Fehr, Efferson, & van Schaik, 2007, cottontop tamarins: Cronin, Kori, & Showdon, 2010, but see also Cronin, Schroeder, Rothwell, Silk, & Snowdon, 2009; Stevens, 2010), and Old World monkeys (long-tailed macaques: Massen, van den Berg, Spruijt, & Sterck, 2010). Bonobos, one of our closest living evolutionary relatives, also showed prosociality in another experimental setting where they could choose to share or not to share food with a conspecific partner (Hare & Kwetuenda, 2010). Surprisingly, chimpanzees have demonstrated “indifference” to the others’ payoff in most of the previous studies using this test paradigm, not discriminating between the prosocial and the selfish options (Brosnan et al., 2009; Jensen, Hare, Call, & Tomasello, 2006; Silk et al., 2005; Vonk et al., 2008; Yamamoto & Tanaka, 2010; but see also Horner, Cartera, Suchak, & de Waal, 2011).
The targeted helping paradigm examines whether animals help other animals in predicaments that they cannot resolve on their own (Fig. 1b). Results to date using this paradigm have been seemingly incompatible with those using the prosocial choice tests (Table 1). Besides humans, chimpanzees are the most helpful animals in the targeted helping paradigm (Melis et al., 2010; Warneken & Tomasello, 2006; Warneken, Hare, Melis, Hanus, & Tomasello, 2007; Yamamoto, Humle, & Tanaka, 2009, 2012). However capuchin monkeys, who have shown prosociality in the prosocial choice paradigm, have demonstrated limited performance in the targeted helping paradigm (Barnes, Martinez, Langer, Hill, & Santos, 2008; Skerry, Sheskin, & Santos, 2011). Although there are a relatively small number of empirical studies in which this paradigm has been used with non-human animal species, some studies have suggested that targeted helping is observed only in limited number of animals, such as some great apes, dolphins, and elephants (de Waal, 2008).
What is the difference between the two test paradigms? What psychological or cognitive abilities are required in each test? In the prosocial choice test, if the subjects are other-regarding, they only need to discriminate the reward distributions between the two options. Therefore, the prosocial choice test is a useful device to evaluate whether animals have other-regarding preferences and whether they can discriminate between the reward distributions for themselves and the partner. Meanwhile, to complete a targeted helping task, animals have to understand their partner’s goals. This paradigm requires of animals more elaborate cognitive abilities in terms of understanding others, i.e., perspective-taking. Capuchin monkeys’ results seem to fit with this explanation. They have shown prosociality in the simpler prosocial choice test (de Waal et al., 2008; Lakshminarayanan & Santos, 2008; Takimoto & Fujita, 2011; Takimoto, Kuroshima, & Fujita, 2010), but not so much in the targeted helping test (Barnes et al., 2008; Skerry et al., 2011). It has been suggested that capuchin monkeys are not so skillful in perspective-taking compared to chimpanzees (Hare, Call, Agnetta, & Tomasello, 2000; Hare, Addessi, Call, Tomasello, & Visalberghi, 2003).
Interestingly, however, chimpanzees have demonstrated the opposite: they have shown prosociality in the targeted helping tests (Melis et al., 2010; Warneken & Tomasello, 2006; Warneken et al., 2007; Yamamoto et al., 2009, 2012), but rarely in the prosocial choice tests (Brosnan et al., 2009; Jensen et al., 2006; Silk et al., 2005; Vonk et al., 2008; Yamamoto & Tanaka, 2010; but see also Horner et al., 2011). One of the most plausible explanations would be that chimpanzees may recognize others’ goals (Call, Hare, Carpenter, & Tomasello, 2004), but may not take them into account unless directly requested to help. Yamamoto et al. (2012) empirically demonstrated that chimpanzees can understand the partner’s goals and flexibly adjust their targeted helping in accordance with what the partner needs. In their experiments, they presented chimpanzees with seven objects, one of which could be used as a tool by the partner who was in an adjacent booth facing one of two different tool-use tasks. In this setting, the helpers selected and transferred an appropriate tool upon request, although not without such a request, when they could visually assess the partner’s situation. Interestingly, when the visual information about the partner’s situation was interrupted, the helpers still tried to respond to the partner’s request (a gesture made by stretching out an arm), but could not select an appropriate tool. These results suggested the following: (1) chimpanzees can understand others’ goals just by visual assessment, (2) in spite of this, chimpanzees seldom help others proactively, and (3) chimpanzees try to help others upon request even when they do not understand the exact goals of others. The importance of the recipients’ request has also been suggested from other empirical studies (Jaeggi, Burkart, & van Schaik, 2010; Melis et al., 2010; Warneken & Tomasello, 2006; Warneken et al., 2007; Yamamoto et al., 2009). In some prosocial choice tests, however, the request did not affect (Vonk et al., 2008) or even decreased (Horner et al., 2011) the chimpanzees’ prosocial choice, which suggests that a request works conditionally and sometimes plays an opposite role, as a harassment which might undermine the prosociality of the partner.
Considering the characteristics of chimpanzee helping, humans appear to be unique in their proactive targeted helping. Sometimes people help others upon observing them in trouble, even without being requested. This may be possible because of humans’ well-developed theory-of-mind abilities, such as understanding others’ desires and shared intentionality. However, as is sometimes seen in real life, unsolicited helping runs the risk of resulting in wasted labor, as the recipient may not need or appreciate the help. Why have humans evolved such a risky helping style? Reputation and indirect reciprocity may have strongly influenced its evolution (Nowak & Sigmund, 2005; Ohtsuki & Iwasa, 2006). Even if the recipient does not appreciate their proactive help, the helper may obtain a good reputation from third-party individuals and then from the society to which they belong. It can be easily argued that in such a society, people behave prosocially even without being solicited (Yamamoto & Tanaka, 2009a). This indirect reciprocity system ensures future benefit for prosocial individuals in the ultimate perspective, which means that people are not necessarily conscious of this system.
Prosociality and Fairness
As stated above, prosociality and fairness are supposed to be closely linked. Especially in humans, the strong preference for fairness, in collaboration with the reputation system, leads to our social norms, which strongly promote prosocial behavior (Fehr & Fischbacher, 2004). Fairness can be behaviorally presented as inequity aversion, which is categorized into two types: advantageous and disadvantageous inequity aversion. The former is a negative response to an inequitable outcome in which an individual earns more than a comparison individual, while the latter means that an individual resists an opposite inequitable situation, in which they receive less than another (Fehr & Schmidt, 1999). People demonstrate both the types of inequity aversion. This means that people are willing to give up some material payoff to benefit others and to move in the direction of more equitable outcomes (advantageous inequity aversion). This human inclination strongly supports proactive prosocial behavior.
Now the question is whether this happens in non-human animals or not. At this moment, there is no empirical evidence for reputation or social norms having a strong effect on prosocial behavior in primates other than humans. Although chimpanzees (Subiaul, Vonk, Okamoto-Barth, & Barth, 2008; Russell, Call, & Dunbar, 2008; but not capuchin monkeys: Brosnan & de Waal, 2009) seem to evaluate other’s behavior, there is no evidence that they adjust their prosocial behavior by recognizing that they are evaluated by others (i.e., audience effect). In addition, although there has been accumulating evidence for a sense of fairness in non-human animals (for review, see Brosnan, 2009, 2011), the animals have normally demonstrated only disadvantageous inequity aversion (Brosnan & de Waal, 2003; Brosnan, Schiff, & de Waal, 2005; Fletcher, 2008; Massen et al., 2011; Neiworth, Johnson, Whillock, Greenberg, Brown, 2009; van Wolkenten, Brosnan, & de Waal, 2007), and there is little evidence for advantageous inequity aversion (Brosnan, Talbot, Ahlgren, Lambeth, & Schapiro, 2010; see also Brosnan, 2012; Christen & Glock, 2012). Therefore, a sense of fairness in non-human animals may not directly promote prosocial behavior in them. Actually, the absence or scarcity of proactive prosocial behavior, especially in chimpanzees, may partly be explained by these facts, i.e., the lack of social norm and advantageous inequity aversion. In this section, we will focus on disadvantageous inequity aversion in non-human animals, and discuss another possible role of this type of sense of fairness and how it influences prosocial behavior.
First of all, there appears to be a considerable phylogenetic link between prosociality and inequity aversion. Chimpanzees and capuchin monkeys have been the most examined. These two species have shown both a greater tendency to reject inequitable outcomes (Brosnan & de Waal, 2003; Brosnan et al., 2005; Brosnan, Talbot, et al., 2010; Fletcher, 2008; van Wolkenten et al., 2007; but see also Jensen et al., 2007a) and greater prosocial tendencies in either the prosocial choice tests or the helping tests than have many other species (de Waal et al., 2008; Lakshminarayanan & Santos, 2008; Melis et al., 2010; Takimoto & Fujita, 2011; Takimoto et al., 2010; Warneken & Tomasello, 2006; Warneken et al., 2007; Yamamoto et al., 2009, 2012). Cottontop tamarins have also demonstrated both traits (prosociality: Cronin et al., 2010, inequity aversion: Neiworth et al., 2009), although there have been contradictory results on prosociality (Cronin and Snowdon, 2008; Cronin et al., 2009; Stevens, 2010). Long-tailed macaques (Macaca fascicularis) have demonstrated a prosocial tendency (Massen et al., 2010), even though they are known as a despotic species with a steep linear hierarchy (Thierry, 2000). Accordingly, they also showed aversion to disadvantageous inequity (Massen et al., 2011). Orangutans (Pongo pygmaeus), which are not known to cooperate to a great degree in the wild, possibly due to their more solitary social organization (van Schaik & van Hooff, 1996), did not show any inequity aversion (Bräuer, Call, & Tomosello, 2006; Brosnan, Flemming, Talbot, Mayo, & Stoinski, 2011). However, the link between prosociality and fairness requires further investigation, because some studies with captive orangutans have suggested that they do show reciprocal cooperation (Dufour, Pele, Neumann, Thierry, & Call, 2008).
We next consider the link between prosociality and fairness from the perspective of its mechanism. Although it is assumed that a sense of fairness is beneficial as a mechanism for stabilizing cooperation (Fehr & Schmidt, 1999), there can be competition in nature between fairness and prosociality (Brosnan, Hauser, et al., 2010). Prosocial behavior may lead to unequal payoffs between the actor and the recipient (Fig. 2). Consider the prosocial choice test. The actor has the choice of either the prosocial or the selfish option. Either option produces the same reward to the actor; therefore, the actor could benefit the partner at no cost. Nevertheless, the actor has to spend labor on this task (e.g., pulling a bar), whereas the partner does nothing. In this situation, selecting the prosocial option results in disadvantageous inequity in terms of effort, although the reward is equal. Animals that are sensitive to disadvantageous inequity have to overcome at least such temporal inequity. In fact, there has been a study suggesting this competition. In an experiment entailing the competing demands of prosociality and equity, capuchin monkeys ceased prosocial behavior when this other-rewarding behavior resulted in considerable disadvantageous inequity in rewards (Brosnan, Hauser, et al., 2010).
One possibility is that non-human animals can recognize reward differences between themselves and others, but are not sensitive to an imbalance in effort. Many studies that suggested inequity aversion in non-human animals (e.g., Brosnan & de Waal, 2003; Brosnan et al., 2005; Brosnan, Talbot, et al., 2010; Fletcher, 2008) have focused on the imbalance in rewards between the target subject and his or her partner, but did not consider the influence of the reward/effort balance on the partner’s side. A smaller number of studies (Massen et al., 2011; van Wolkenten et al., 2007) manipulated the amount of effort required of the targeted subjects who would get a lesser reward and found that an increased amount of effort increased the subjects’ aversion to the reward inequity; however, this result can be interpreted as a sign that the subjects responded just to their own increased amount of labor, and that this effort-effect might not be based on comparisons with the partner. So far, there has been no empirical study that shows non-human animals are sensitive to inequity in effort between themselves and their partner (cf. Fontenot, Watson, Roberts, & Miller, 2007). Therefore, how animals perceive imbalance and whether or not the animals really do the two-by-two comparison with respect to effort remains to be resolved in future studies.
Nevertheless, some have suggested that capuchin monkeys, although not, so far, chimpanzees (Melis, Schneider, & Tomasello, 2011), are sensitive to the amount of others’ labor at least in cooperation tasks. De Waal and Berger (2000) found that capuchin monkeys shared more food reward with the partner when the reward was obtained by cooperation with the partner than when they obtained the reward by themselves. Moreover, Takimoto and Fujita (2011) showed that capuchin monkeys did not discriminate between prosocial and selfish choices when the partner did not need to help, but actively tried to benefit their partner when the partner did help. Food sharing in de Waal and Berger (2000) could have been a by-product of increased proximity between the monkeys rather than an intention of the operator. However, this account fails to apply to Takimoto and Fujita (2011) as the operator monkeys had to make a dichotomous choice, and there was no increased proximity between the operator and the recipient. These results suggest that non-human animals are equipped with sensitivity to the amount of others’ efforts as well as sensitivity to the reward distribution.
Considering competition between a sense of fairness and prosociality, inequity aversion may not actively “promote” prosociality during the first stage, but may “stabilize” cooperation, by keeping the balance between giving and receiving prosocial behavior. There can be two mechanisms to stabilize cooperation: one with a negative response directly aimed at free-riders, such as punishment that leads free-riders to amend their behavior, and the other is one that involves an evaluation of the partner, enabling animals to avoid free-riders and choose appropriate partners for cooperative interactions. In non-human animal studies, there has been limited evidence supporting the idea that punishment maintains cooperation by changing free-riders into valuable cooperators. Even in humans it has been reported that punishment leads to lower payoffs in repeated public goods games (Rand, Dreber, Ellingsen, Fudenberg, & Nowak, 2009). Thus, we can question whether costly punishment is an ideal force for promoting cooperation, at least in all situations. It is almost certain that non-human animals, especially chimpanzees, have a revenge system, i.e., negative reciprocity (de Waal & Luttrell, 1988; Jensen et al., 2007b). However, such a negative response to inequity often results in the corruption of reciprocal cooperation (Yamamoto & Tanaka, 2009b, c), rather than promoting cooperation (although see Raihani, Grutter, & Bshary, 2010; Raihani & McAuliffe, 2012).
It may be more plausible to assume that instead, a sense of fairness stabilizes cooperation through the mechanism of the evaluation of the partner (Fehr & Schmidt, 1999; Brosnan, 2006, 2011). The ability to recognize situations in which one is receiving a lesser outcome than a partner in a cooperative interaction may encourage individuals to switch to a new partner. If the new partner is more equitable, this ultimately functions to increase the individuals’ payoffs and ensures proper benefits for both. The negative response to inequity can also function as a commitment device (Frank, 1988; Yamagishi et al., 2009). It can be an indicator of an individual’s fairness, whereby a good reputation as a good cooperative partner is obtained (Frank, 1988). Similarly, refusing absolute gains which are relatively unequal may send a signal to potential partners that they cannot get away with such behavior with you, perhaps increasing future gains (Yamagishi et al., 2009). This theory can possibly be applied to non-human animals. Melis, Hare, and Tomasello (2006) reported that chimpanzees facing a collaboration task recruited the more collaborative and effective of two potential partners on the basis of their experience with each of them on a previous day. This suggests that non-human animals, or at least chimpanzees, can evaluate others’ cooperative tendencies and appropriately choose the cooperative partner, which leads to reciprocal interaction (Melis, Hare, & Tomasello, 2008). Although reciprocal cooperation did not last long when the pairs of chimpanzees were fixed in experimental settings (Yamamoto & Tanaka, 2009b, c), it has been suggested that reciprocal interaction is balanced between individuals in the wild (Gomez, Mundry, & Boesch, 2008; Mitani, 2006; Mitani & Watts, 2001). This may be achieved due to their fission–fusion society that allows individuals to flexibly change their partner in social interactions.
As stated above, fairness and cooperation seem to be closely linked phylogenetically, and possibly coevolved (Brosnan, 2011). However, the mechanism remains a theoretical concept supported by inadequate collateral evidence and should be empirically examined in the future studies. If a sense of fairness is actually a useful device for detecting free-riders and choosing partners, this mechanism ensures a deterrent for defection; nevertheless, it cannot actively promote prosocial behavior. In concluding this section, a sense of fairness may stabilize cooperation, but it is unlikely to play a promoter role. To start cooperative relationships by initiating prosocial behavior, there has to be a mechanism to overcome inequity aversion during the first stage. How do animals realize this, and what actively promotes the commencement of prosocial behavior?
Prosociality and Empathy
It is quite plausible that a sense of fairness is closely related to prosocial behavior. Moreover, it may play a role as a stabilizer of prosocial behavior, albeit not a promoter. Therefore, there is a need to identify another psychological mechanism that acts as a promoter of prosocial behavior. Empathy seems to be the best candidate for this role (de Waal, 2008).
Empathy is the capacity to (1) be affected by and share the emotional state of another (e.g., emotional contagion), (2) assess the reasons for the other’s state, and/or (3) identify with the other, adopting his or her perspective (de Waal, 2008; de Waal & Suchak, 2010). This definition of empathy does not necessarily require developed cognitive abilities. Psychologists sometimes speak of empathy only when it involves perspective-taking and are skeptical about the existence of empathy in non-human animals (Hauser, 2000; Povinelli, 1998). However, de Waal (2008) emphasized that at its most basic, empathy can be defined as emotional sensitivity to others or emotional contagion (Hatfield et al., 1994). According to de Waal (2008), perception of the emotional state of another automatically activates shared representations, causing a matching emotional state in the observer. He also proposed a model in which empathy is cognitively multilayered: from state-matching (e.g., emotional contagion) to sympathetic concern (e.g., consolation) and perspective-taking (e.g., targeted helping) with increasing cognitive capacities (de Waal, 2008, 2012).
How does empathy promote prosocial behavior? Previously, de Waal et al. (de Waal, 2008; de Waal & Suchak, 2010) pointed out that sympathetic concern occurs when emotional contagion is combined with the appraisal of the other’s situation, leading to attempts to understand the cause of the other’s emotions. This other-oriented emotional activation might be seen in consoling and assisting others against aggression in the wild. When the ability to understand others (i.e., perspective-taking) is added, it enables animals to perform targeted helping, i.e., help and care based on a cognitive appreciation of the other’s specific need, or situation. This explains the mechanism by which empathy may cause prosocial behavior. In addition, empathy may also work as a feedback system based on the consequences of prosocial behavior. If animals can be affected by the positive emotion of another that they have helped, this emotional contagion will work as a positive feedback to promote future prosocial behavior. People sometimes worry about whether their helping is actually helpful to others and also get upset when their offer of help is refused. Thus, in humans, expression of gratitude is important, even if this is not an actual return benefit. Although this has not been fully examined in non-human animals (but see Bonnie & de Waal, 2004), chimpanzee helpers in the targeted helping test sometimes closely watched the partner getting a reward with a tool that they donated (Yamamoto et al., unpublished data; cf. see supplemental videos in Yamamoto et al., 2009, 2012). They did not seem to be interested in the reward itself in the partner’s hand, as they often demonstrated no request for it. Thus, even though they received no return benefit (e.g., there was no sharing the juice reward), they continued to help the partner.
Therefore, according to this explanation, prosocial behavior may be solicited by empathy in situations in which animals are able to recognize another’s desire easily. Returning to the laboratory findings, we see that in the prosocial choice test, the partner’s need was obvious in the simplest setup: for the prosocial option, food was set in front of the partner, and their own and the partner’s desire for food was the same. Takimoto and Fujita (2011) reported that capuchin monkeys participating as a “partner” often made a request to the subject, or expressed a desire for the food by reaching out for it. Therefore, if we can assume the animals are equipped with sensitivity to another’s emotion, prosocial behavior in this choice test could naturally occur in the animals.
Now the question is why chimpanzees did not proactively help others even when they understood exactly the partner’s need (e.g., Yamamoto et al., 2012). In the targeted help tests, chimpanzees understood what the partner needed and appropriately selected and donated a tool to the partner so that they could get a reward using it. This study also suggested that the chimpanzees could only understand the partner’s goal by visual assessment, but not by direct vocal and gestural communication between them. In spite of this, they did not help the partner proactively, but only after being requested. Why did prosociality not occur automatically following the chimpanzees’ understanding of the needs of their partners?
Here, we may have to separately consider empathy and the understanding of others’ goal. de Waal stated (2008 p. 285) that “Perspective-taking by itself is, of course, hardly empathy: It is so only in combination with emotional engagement”. Continuing this line of discussion, cognitively developed species do not necessarily show high levels of empathy or prosociality.
We would like to advance this discussion by hypothesizing that cognitively developed animals have to be equipped with some modulatory mechanism to deal with empathy-based prosociality. If animals automatically responded to every desire of others with prosocial behaviors, they would be exhausted from helping. Such individuals could also be easily exploited by free-riders, which would result in the extinction of prosociality. Therefore, animals that have the capacity to put themselves in the other’s “shoes” and can easily recognize the other’s desire paradoxically have to limit their helping to some extent. As a result, in animals that are skilled at perspective-taking, their inhibitory mechanism should also be strong, and therefore some additional triggers should be required for eliciting prosocial behavior. This hypothesis fits the results for chimpanzees. Chimpanzees may recognize others’ goals and needs, but may not take them into account unless directly requested for help.
How do animals limit their prosociality? One strategy is to refrain from proactively helping, as the chimpanzees do in the tests described above. Social relationships may also have a considerable influence on this process. The closest relationship is the mother–infant relationship, which can be considered as the core unit of a social group (Hirata, 2009; Matsuzawa, Tomonaga, & Tanaka, 2006). Prosocial behavior occurs most often between the mother and infant (Hirata, 2009), although request is still important even in this closest bond in chimpanzees (Ueno & Matsuzawa, 2004; Yamamoto et al., 2009; Yamamoto & Tanaka, 2009c, 2010); other kin, friends, old acquaintances, and in-group members follow this in frequency. In capuchin monkeys, it was reported that prosocial tendencies increased with social closeness, being lowest toward strangers and highest toward kin (de Waal et al., 2008). In long-tailed macaques, Massen et al. found a strong effect of kin-ship on prosocial behavior (Massen et al., 2010), although they did not find an effect of relationship quality among non-kin (Massen et al., 2010, 2011). Competition may also serve as an inhibitor of prosociality. It is suggested that chimpanzees demonstrate cognitively sophisticated abilities in experiments using competition paradigms, as compared to cooperation paradigms (Hare, 2001; Hare & Tomasello, 2004). Therefore, chimpanzees seem to be fine-tuned to competition, in which naturally the automatic arousal of prosociality should be minimal.
Another way that animals limit prosociality is through fairness. Disadvantageous inequity aversion serves to stop the unnecessarily excessive expression of prosociality. When others are getting an equal or greater benefit, there is no need to help them. A sense of fairness plays a role in this evaluation. A sense of fairness also helps animals choose an appropriate partner to whom they should direct their prosociality. In humans, research on neural activity suggests that empathic brain responses to another person’s pain are modulated by perceived fairness of the other person (Singer et al., 2006). In Singer et al.’s (2006) experiments, empathy-related activation was observed when a fair, likeable player was in pain. However, men, but not women, showed an absence of such empathic brain response when seeing an unfair player in pain. The link between fairness and empathy will be discussed again in the section “Fairness and empathy.”
In conclusion for this section, empathy can serve as a promoter of prosocial behavior, but does not possess in itself mechanisms for stabilizing cooperation (i.e., free-rider avoidance). Therefore, animals equipped with empathy need a braking system to avoid being exploited by free-riders. We suggest that empathy and fairness may work interactively with each other to maintain the balance of prosociality. We will discuss this in detail in the next section.
Fairness and Empathy
Here is a brief review of our points. Empathy promotes but does not stabilize prosocial behavior, whereas a sense of fairness stabilizes but does not promote. These two psychological mechanisms can be likened to a car’s accelerator and brake. To drive a car (prosociality and cooperation), both an accelerator (empathy) and a brake (a sense of fairness) are necessary, and balance between the two is important. A brake of course, cannot start a car, and an accelerator cannot stop a car from crashing.
Emerging data support the hypothesis that a sense of fairness and empathy coevolved in conjunction with prosociality. Comparative studies indicate that species that respond to inequitable outcomes also demonstrate empathic prosocial behavior (Table 1). Although we are limited to data from only a few species, studies on chimpanzees and capuchin monkeys have shown disadvantageous inequity aversion in these species (chimpanzees: Brosnan et al., 2005; Brosnan, Talbot, et al., 2010, capuchin monkeys: Brosnan & de Waal, 2003; Fletcher, 2008; van Wolkenten et al., 2007). They are also known to be other-regarding in some ways (chimpanzees: Horner et al., 2011; Melis et al., 2010; Warneken & Tomasello, 2006; Warneken et al., 2007; Yamamoto et al., 2009; 2012, capuchin monkeys: de Waal et al., 2008; Lakshminarayanan & Santos, 2008; Takimoto & Fujita, 2011; Takimoto et al., 2010). Bonobos have been considered to be prosocial and more cooperative than chimpanzees (Hare & Kwetuenda, 2010; Hare, Melis, Woods, Hastings, & Wrangham, 2007; but see also Jaeggi, Stevens, & van Schaik, 2010), and in a study of direct comparison with chimpanzees and orangutans, they have demonstrated the highest difference in the rejection rate between the inequity (approximately 20 % rejection) and equity conditions (approximately 10 % rejection), although this difference was not significant (possibly due to the small sample size; see the discussion in Brosnan, 2011 referring to Bräuer et al., 2009; see also Bräuer & Hanus, 2012). Recently, long-tailed macaques (who are a despotic species) have demonstrated both empathic prosociality (Massen et al., 2010) and inequity aversion (Massen et al., 2012). We cannot directly examine empathy itself from behavioral data; however, in experimental paradigms (prosocial choice tests and targeted helping tests) animals could not expect any return benefit, and therefore the psychological motivation could be interpreted as being driven by empathy (de Waal, 2008).
The hypothesis that a sense of fairness and empathy coevolved is also supported by data on the absence of prosocial behavior. Among the New World monkeys, squirrel monkeys, which cooperate in only limited situations (Boinski, 1987), did not respond negatively to inequity (Talbot, Freeman, Williams, & Brosnan, 2011). Data on cottontop tamarins are not fully consistent. In some studies, cottontop tamarins demonstrated sympathetic concerns (Cronin et al., 2010); however, in others they did not show other-regarding behavior in the prosocial choice tests (Cronin et al., 2009; Stevens, 2010). Their tendency of inequity aversion is also still debated (cf. Neiworth et al., 2009; see also Price & Brosnan, 2012). Considering that cottontop tamarins are cooperative breeders and have demonstrated intense levels of cooperation (Cronin et al., 2005; Cronin & Snowdon, 2008), we can expect that they are equipped with these two psychological traits; however, at this moment, we just point out the necessity of further investigation with this species. Common marmosets, another cooperative-breeding New World monkey, have shown prosociality, choosing the prosocial option more often when the partner was present than absent (Burkart et al., 2007). Unfortunately, at this moment, data on inequity aversion in this species are not available. Again further studies are needed.
Both a sense of fairness and empathy might have evolved on the basis of preference for similarity with others. Humans, from a very early age, are known to show preference to individuals who are similar to themselves. Sanefuji, Ohgami, and Hashiya (2006) presented 6- and 9-month-old babies with movies and pictures of 6-, 9-, and 12-month olds. The results revealed that 9-month olds showed peer preference when they saw movies of infants and that 6- and 9-month olds preferred static images of same-age infants. This study suggests that humans inherently have a sensitivity to similarities and differences between others and themselves, and that they feel familiarity with those who have similar traits. Such sensitivity can be considered as a basis for fairness and empathy. Similarity between individuals seems to be important in affiliative interactions among non-human primates, too. Capuchin monkeys affiliate preferably with human experimenters who imitated their behavior (Pankner, Suomi, Visalberghi, & Ferrari, 2009). Primates sometimes intentionally adjust their power balance between individuals. For example, analysis of play behavior in primates indicates that physically stronger, or socially dominant participants tend to inhibit their power and control playful interactions with weaker or subordinate individuals (self-handicapping: Fagan, 1981; Shimada, 2006). As a result, actual differences in relative ranks, or power among participants, do not seem to exist during social play, which leads to maintaining this affiliative interaction (but see also van Leeuwen, Zimmermann, & Davila Ross, 2010: gorillas seem to try to maintain their competitive advantage during play fights).
Peer preference in non-human animals can also be easily seen in their in-group biases in friendly social interactions. They restrict cooperative behavior to familiar group members. In the experiments that have examined responses to inequity, it has been suggested that chimpanzees, but not long-tailed macaques (Massen et al., 2012), show higher tolerance to inequity between long-term associates, in comparison to recently introduced individuals (Brosnan et al., 2005). Thus, inequity aversion is regulated by the social relationship between individuals. Social closeness also affects empathy, although in the opposite direction, by enhancing empathy. In prosocial choice tests, prosocial tendencies in capuchin monkeys increased with social closeness, being lowest toward strangers and highest toward kin (de Waal et al., 2008). In chimpanzees, mutual request behaviors were observed in mother–infant pairs more frequently than in non-kin adult pairs, leading to more frequent and mutual helping between mothers and infants (Yamamoto et al., 2009). As mentioned in the section “Prosociality and empathy,” a sense of fairness and empathy are closely related: when feeling inequity, men did not feel empathy for the unfair person in pain. For the emergence of prosociality, the balance between a sense of fairness and empathy is important, and this seems to be regulated by the social relationship with others.
In short, we postulate that a sense of fairness and empathy emerged from the same principle and work interactively and differently for soliciting and maintaining prosocial behavior. Although empathy, or at least its perception–action mechanism (Preston & de Waal, 2002), is an automatic response that does not require conscious effort, a sense of fairness may require a higher level of cognition, such as comparing the reward/effort balance with others (Fehr & Schmidt, 1999). Therefore, emotional contagion, the first step of empathy, may be found in various mammals and birds (de Waal, 2008), but the existence of a sense of fairness may be restricted to several more cognitively-developed species (including non-primates; see Range, Leitner, & Virányi, 2012; Horowitz, 2012; Pierce & Bekoff, 2012). We hypothesize that if both a sense of fairness and empathy are important for promoting and stabilizing prosocial behavior, empathy (emotional contagion) alone may not be related to the prevalence of prosociality, in the absence of a sense of fairness, which may be considered to be the braking mechanism. For example, mirror neurons seem to compose the core part of empathy, such as the perception–action mechanism, but may not warrant sympathetic concern in animals (Preston & de Waal, 2002). Thus, the existence of mirror neurons itself does not directly lead the existence of prosociality in a species.
Conclusion and Future Direction
In this article, we have discussed prosocial behavior and cooperation in relation to their possible underlying mechanisms, a sense of fairness and empathy. We then proposed a hypothesis of the coevolution of a sense of fairness and empathy and suggest that they function as a promoter and a stabilizer of prosocial behavior, respectively. Of course, not all prosocial behavior requires empathy (de Waal, 2008), and the propensity to engage in other-regarding behavior is not determined exclusively by whether an animal is equipped with such empathic resonance mechanisms, but by many other factors as well (Singer, 2009). Keeping this in mind, we simplified our discussion by focusing on these two mechanisms and the triadic relationship.
Figure 3 exemplifies how prosocial behavior and cooperation are related to a sense of fairness, empathy, and other relevant factors. We suggest that starting from the preference for similarity, perhaps through the avoidance of heterogeneity, psychological mechanisms for inequity aversion, i.e., a sense of fairness, emerged. This may have developed into social norms when animals acquired a reputation system and its required cognitive abilities, such as language and an understanding of triadic interactions (self, partner, and third-party), which can be found specifically in humans (Tomonaga et al., 2004). Although social norms may promote prosocial behavior directly, a sense of fairness, represented as disadvantageous inequity aversion, functions to stabilize prosocial behavior through the evaluation of others’ fairness, and partner choice. The other line of mechanisms, i.e., empathy, also starts from psychological traits that prefer similarity with others, going through simple emotional contagion and leading to sympathetic concern. Although the ability to understand others, such as perspective-taking, is not necessarily connected to prosocial behavior, empathy, and especially sympathetic concern, function to promote prosocial behavior directly.
We have reviewed a number of comparative studies conducted with non-human primates that at least partially support our hypothesis. Unfortunately, however, there are still not enough data to conclusively support, or reject, our hypothesis. First of all, controversy still continues as to whether a sense of fairness exists in non-human animals (Bräuer et al., 2009; Henrich, 2004; Silberberg, Crescimbene, Addessi, Anderson, & Visalberghi, 2009; see also Christen & Glock, 2012). To counter this skepticism, we suggest that researchers have to focus more on the balance between reward and effort (see also Skitka, 2012). Previously studies with non-human animals have focused mainly on reward inequity, almost ignoring the effort imbalance. Furthermore, we also have to consider individual differences. Who is concerned about fairness? It is possible that individual A may feel inequity to a certain payoff that individual B may consider fair? Social status, physical abilities, and many other factors might affect a sense of fairness. When discussing a sense of fairness in this article, we focused on disadvantageous inequity aversion because there has been little evidence for advantageous inequity aversion in non-human animals (Brosnan, Talbot, et al., 2010). As mentioned above (see the section “Prosociality and fairness”), advantageous inequity aversion is directly linked to prosocial behavior, and some researchers have pointed out the possibility that human prosocial behavior, as observed in economic game paradigms, may well be based on fairness motives rather than empathic motives (Singer, 2009). The evolution of advantageous inequity aversion is still mysterious and worth further investigation.
As for empathy, many issues also remain to be investigated. Preston and de Waal (2002) proposed the perception–action model in which empathy is automatically aroused upon observing others in some emotional states. However, considering the observed empathy–fairness intervention in neural activities, which is found in men who did not feel empathy for the unfair person in pain (Singer et al., 2006), it is plausible that empathy is not always a mere automatic consequence of the passive observation of emotional cues in others, but rather subject to an appraisal of the situation and modulation (de Vignemont & Singer, 2006; Singer, 2009). The interaction between fairness and empathy recently received attention in empirical studies with non-human animals (e.g., de Waal et al., 2008), and further developments are expected.
In human prosociality, we can detect the influence of a sense of fairness and empathy, as well as their interaction, and it seems that human psychological tendencies are more biased toward prosociality, which allows for our highly cooperative society. From the viewpoint of cognitive mechanisms, this seems to be linked to what appear to be human-unique traits, including advantageous inequity aversion, social norms, and sophisticated abilities to understand others’ desires, although perspective-taking itself should be carefully discussed apart from sympathetic concern. In addition to searching for the evolutional basis for these cognitive mechanisms, we would also like to understand individual, sex, cultural, and/or developmental differences in human prosociality, fairness, and empathy. As the literature on these topics has begun to develop only recently (e.g., Singer et al., 2006; Svetlova, Nichols, & Brownell, 2010), there is not yet enough evidence to draw firm conclusions. A better understanding of the nature, culture, and development of the promoting and modulating factors of prosociality will help us to understand the complex interplay of conditions determining prosocial behavior.
Acknowledgments
We thank Dr. Sarah Brosnan, the editor of this pair of special issues, and the anonymous reviewers for their thoughtful comments. This study was financially supported by the Japan Society for the Promotion of Science Grants #22800034 (to S.Y.) and #20220004 (to A.T.), and by the Ministry of Education, Culture, Sports, Science & Technology in Japan #24000001 (to Tetsuro Matsuzawa).
References
- Barnes, J. L., Martinez, M., Langer, M., Hill, T., & Santos, L. R. (2008). Helping behavior and regard for others in capuchin monkeys (Cebus apella): An evolutionary perspective on altruism. Biology Letters, 4(6), 638–640.
- Boesch, C., & Boesch, H. (1989). Hunting behavior of wild chimpanzees in the Tai National Park. American Journal of Physical Anthropology, 78, 547–573.
- Boinski, S. (1987). Mating patterns in squirrel monkeys (Saimiri oestedi): Implications for sexual dimorphism. Behavioral Ecology and Sociobiology, 21, 13–21.
- Bonnie, K., & de Waal, F. B. M. (2004). Primate social reciprocity and the origin of gratitude. In R. Emmons & M. McCullough (Eds.), The psychology of gratitude (pp. 213–229). Oxford: Oxford University Press.
- Bra¨uer, J., Call, J., & Tomasello, M. (2009). Are apes inequity averse? New data on the token-exchange paradigm. American Journal of Primatology, 7, 175–181.
- Bra¨uer, J., Call, J., & Tomosello, M. (2006). Are apes really inequity averse? Proceedings of the Royal Society, London, B, Biological Science, 273, 3123–3128.
- Bra¨uer, J., & Hanus, D. (2012). Fairness in non-human primates? Social Justice Research, 25(3). doi: 10.1007/s11211-012-0159-6.
- Brosnan, S. F. (2006). Nonhuman species' reactions to inequity and their implications for fairness. Social Justice Research, 19, 153–185.
- Brosnan, S. F. (2009). Responses to inequity in non-human primates. In P. W. Glimcher, C. F. Camerer, E. Fehr, & R. A. Poldrack (Eds.), Neuroeconomics (pp. 285–301). New York: Academic Press.
- Brosnan, S. F. (2011). A hypothesis of the co-evolution of cooperation and responses to inequity. Frontiers in Neuroscience, 5, 43. doi:10.3389/fnins.2011.00043.
- Brosnan, S. F. (2012). Introduction to ‘‘Justice in Animals''. Social Justice Research, 25(2), 109–121.
- Brosnan, S. F., & de Waal, F. B. M. (2003). Monkeys reject unequal pay. Nature, 425, 297–299.
- Brosnan, S. F., & de Waal, F. B. M. (2009). Capuchin monkeys tolerate intermittent unreliability in human experimenters. International Journal of Primatology, 30, 663–674.
- Brosnan, S. F., Flemming, T., Talbot, C. F., Mayo, L., & Stoinski, T. (2011). Orangutans (Pongo pygmaeus) do not form expectations based on their partner's outcomes. Folia Primatologica, 82(1), 56–70.
- Brosnan, S. F., Hauser, D., Leimgruber, K., Xiao, E., Chen, T., & de Waal, F. B. M. (2010). Competing demands of prosociality and equity in monkeys. Evolution and Human Behavior, 41, 279–288.
- Brosnan, S. F., Schiff, H. C., & de Waal, F. B. M. (2005). Tolerance for inequity may increase with social closeness in chimpanzees. Proceedings of the Royal Society, London, B, Biological Science, 1560, 253–258.
- Brosnan, S. F., Silk, J. B., Henrich, J., Mareno, M. C., Lambeth, S. P., & Schapiro, S. J. (2009). Chimpanzees (Pan troglodytes) do not develop contingent reciprocity in an experimental task. Animal Cognition, 12, 587–597.
- Brosnan, S. F., Talbot, C., Ahlgren, M., Lambeth, S. P., & Schapiro, S. J. (2010). Mechanisms underlying responses to inequitable outcomes in chimpanzees, Pan troglodytes. Animal Behaviour, 79, 1229–1237.
- Burkart, J., Fehr, E., Efferson, C., & van Schaik, C. P. (2007). Other-regarding preferences in a nonhuman primate: Common marmosets provision food altruistically. Proceedings of the National Academy of Sciences, USA, 104, 19764–19766.
- Busse, C. D. (1978). Do chimpanzees hunt cooperatively? American Naturalist, 112, 767–770.
- Call, J., Hare, B., Carpenter, M., & Tomasello, M. (2004). ‘Unwilling' versus ‘unable': Chimpanzees' understanding of human intentional action. Developmental Science, 7, 488–498.
- Chalmeau, R. (1994). Do chimpanzees cooperate in a leaning task? Primates, 35, 385–392.
- Christen, M., & Glock, H.-J. (2012). The (limited) space for justice in social animals. Social Justice Research, 25(3). doi:10.1007/s11211-012-0163-x.
- Crawford, M. P. (1937). The cooperative solving of problems by young chimpanzees. Comparative Psychology Monographs, 14, 1–88.
- Cronin, K. A., Kori, K. E. S., & Showdon, C. T. (2010). Prosocial behavior emerges independent of reciprocity in cottontop tamarins. Proceedings of the Royal Society, London, B, Biological Science, 1701, 3845–3851.
- Cronin, K. A., Kurian, A. V., & Snowdon, C. T. (2005). Cooperative problem solving in a cooperatively breeding primates. Animal Behaviour, 75, 245–257.
- Cronin, K. A., Schroeder, K. K. E., Rothwell, E. S., Silk, J. B., & Snowdon, C. T. (2009). Cooperatively breeding cottontop tamarins (Saguinus oedipus) do not donate rewards to their long-term mates. Journal of Comparative Psychology, 123, 231–241.
- Cronin, K. A., & Snowdon, C. T. (2008). The effects of unequal reward distributions on cooperative problem solving by cotton-top tamarins (Saguinus oedipus). Animal Behaviour, 75, 245–257.
- de Vignemont, F., & Singer, T. (2006). The empathic brain: How, when and why? Trends in Cognitive Sciences, 10, 435–441.
- de Waal, F. B. M. (2008). Putting the altruism back into altruism: The evolution of empathy. Annual Review of Psychology, 59, 279–300.
- de Waal, F. B. M. (2012). The antiquity of empathy. Nature, 336, 874–876.
- de Waal, F. B. M., & Berger, M. L. (2000). Payment for labour in monkeys. Nature, 404, 563.
- de Waal, F. B. M., Leimgruber, K., & Greenberg, A. R. (2008). Giving is self-rewarding for monkeys. Proceedings of the National Academy of Sciences, USA, 105, 13685–13689.
- de Waal, F. B. M., & Luttrell, L. M. (1988). Mechanisms of social reciprocity in three primate species: Symmetrical relationship characteristics or cognition? Ethology and Sociobiology, 9, 101–118.
- de Waal, F. B. M., & Suchak, M. (2010). Prosocial primates: Selfish and unselfish motivations. Philosophical Transactions of the Royal Society, B, 365, 2711–2722.
- Dufour, V., Pele, M., Neumann, M., Thierry, B., & Call, J. (2008). Calculated reciprocity after all: Computation behind token transfers in orangutans. Biology Letters, 5, 172–175.
- Dugatkin, L. A. (1997). Cooperation among animals: An evolutionary perspective. Oxford: Oxford University Press.
- Eisenberg, N., & Mussen, P. H. (1989). The roots of prosocial behavior in children. Cambridge: Cambridge University Press.
- Fagan, R. (1981). Animal play behavior. Oxford: Oxford University Press.
- Fehr, E., & Fischbacher, U. (2004). Social norms and human cooperation. Trends in Cognitive Sciences, 8, 185–190.
- Fehr, E., & Schmidt, K. M. (1999). A theory of fairness, competition, and cooperation. The Quarterly Journal of Economics, 114, 817–868.
- Fletcher, G. E. (2008). Attending to the outcome of others: Disadvantageous inequity aversion in male capuchin monkeys (Cebus apella). American Journal of Primatology, 70, 901–905.
- Fontenot, M. B., Watson, S. L., Roberts, K. A., & Miller, R. W. (2007). Effects of food preferences on token exchange and behavioural responses to inequality in tufted capuchin monkeys, Cebus apella. Animal Behaviour, 74, 487–496.
- Frank, R. H. (1988). Passions within reason: The strategic role of the emotion. New York: W.W. Norton & Co.
- Gomez, C. M., Mundry, R., & Boesch, C. (2009). Long-term reciprocation of grooming in wild West African chimpanzees. Proceedings of the Royal Society, London, B, Biological Science, 276, 699–706.
- Hamilton, W. D. (1964). The genetical evolution of social behavior. 1 and 2. Journal of Theoretical Biology, 7, 1–52.
- Hare, B. (2001). Can competitive paradigms increase the validity of social cognitive experiments on primates? Animal Cognition, 4, 269–280.
- Hare, B., Addessi, E., Call, J., Tomasello, M., & Visalberghi, E. (2003). Do capuchin monkeys, Cebus paella, know what conspecifics do and do not see? Animal Behaviour, 65, 131–142.
- Hare, B., Call, J., Agnetta, B., & Tomasello, M. (2000). Chimpanzees know what conspecifics do and do not see. Animal Behaviour, 59, 771–785.
- Hare, B., & Kwetuenda, S. (2010). Bonobos voluntarily share their own food with others. Current Biology, 20, R1–R2.
- Hare, B., Melis, A. P., Woods, V., Hastings, S., & Wrangham, R. (2007). Tolerance allows bonobos to outperform chimpanzees on a cooperative task. Current Biology, 17, 1–6.
- Hare, B., & Tomasello, M. (2004). Chimpanzees are more skilful in competitive than in cooperative cognitive tasks. Animal Behaviour, 68, 571–581.
- Hatfield, E., Cacioppo, J. T., & Rapson, R. L. (1994). Emotional contagion. Cambridge, England: Cambridge University Press.
- Hauser, M. D. (2000). Wild Minds: What animals really think?. New York: Holt.
- Henrich, J. (2004). Inequity aversion in capuchins? Nature, 428, 139.
- Hirata, S. (2009). Chimpanzee social intelligence: Selfishness, altruism, and the mother–infant bond. Primates, 50, 3–11.
- Hirata, S., & Fuwa, K. (2007). Chimpanzees (Pan troglodytes) learn to act with other individuals in a cooperative task. Primates, 48, 13–21.
- Horner, V., Cartera, J. D., Suchak, M., & de Waal, F. B. M. (2011). Spontaneous prosocial choice by chimpanzees. Proceedings of the National Academy of Sciences, USA, 108, 13847–13851.
- Horowitz, A. (2012). Fair is fine, but more is better: Limits to inequity aversion in the domestic dog. Social Justice Research, 25, 195–212.
- Jaeggi, A. V., Burkart, J. M., & van Schaik, C. P. (2010a). On the psychology of cooperation in humans and other primates: Combining the natural history and experimental evidence of prosociality. Philosophical Transactions of The Royal Society B, 365, 2723–2735.
- Jaeggi, A. V., Stevens, J. M. G., & van Schaik, C. P. (2010b). Tolerant food sharing and reciprocity is precluded by despotism among bonobos but not chimpanzees. American Journal of Physical Anthropology, 143, 41–51.
- Jensen, K., Call, J., & Tomasello, M. (2007a). Chimpanzees are rational maximizers in an ultimatum game. Science, 318, 107–109.
- Jensen, K., Call, J., & Tomasello, M. (2007b). Chimpanzees are vengeful but not spiteful. Proceedings of the National Academy of Sciences, USA, 104, 13046–13050.
- Jensen, K., Hare, B., Call, J., & Tomasello, M. (2006). What's in it for me? Self-regard precludes altruism and spite in chimpanzees. Proceedings of the Royal Society, London, B, Biological Science, 273, 1013–1021.
- Kappeler, P. M., & van Schaik, C. P. (Eds.). (2006). Cooperation in primates and humans: Mechanisms and evolution. Berlin: Springer.
- Krebs, J. R., & Davies, N. B. (1987). An introducton to behavioural ecology (2nd ed.). Oxford: Blackwell Scientific Publications.
- Lakshminarayanan, V. R., & Santos, L. R. (2008). Capuchin monkeys are sensitive to others' welfare. Current Biology, 18, 999–1000.
- Massen, J. J. M., Luyten, I. J. A. F., Spruijt, B. M., & Sterck, E. H. M. (2011). Benefiting friends or dominants: Prosocial choices mainly depend on rank position in long-tailed macaques (Macaca fascicularis). Primates, 52, 237–247.
- Massen, J. J. M., van den Berg, L. M., Spruijt, B. M., & Sterck, E. H. M. (2010). Generous leaders and selfish underdogs: Pro-sociality in despotic macaques. PLoS ONE, 5, e9734.
- Massen, J. J. M., van den Berg, L. M., Spruijt, B. M., & Sterck, E. H. M. (2012). Inequity aversion in relation to effort and relationship quality in long-tailed macaques (Macaca fascicularis). American Journal of Primatology, 74, 145–156.
- Matsuzawa, T., Tomonaga, M., & Tanaka, M. (Eds.). (2006). Cognitive development in chimpanzees. Tokyo: Springer.
- Maynard-Smith, J. (1982). Evolution and the theory of games. Cambridge: Cambridge University Press.
- Melis, A. P., Hare, B., & Tomasello, M. (2006). Chimpanzees recruit the best collaborators. Science, 311, 1297–1300.
- Melis, A. P., Hare, B., & Tomasello, M. (2008). Do chimpanzees reciprocate received favors? Animal Behaviour, 76, 951–962.
- Melis, A. P., Schneider, A., & Tomasello, M. (2011). Chimpanzees share food in the same way after collaborative and individual food acquisition. Animal Behaviour, 82, 485–493.
- Melis, A. P., & Semmann, D. (2010). How is human cooperation different? Philosophical Transactions of the Royal Society B, 365, 2663–2674.
- Melis, A. P., Warneken, F., Jensen, K., Schneider, A. C., Call, J., & Tomasello, M. (2010). Chimpanzees help conspecifics obtain food and non-food items. Proceedings of the Royal Society, London, B, Biological Science, 278, 1405–1413.
- Mitani, J. C. (2006). Reciprocal exchange in chimpanzees and other primates. In P. M. Kappeler & C. P. van Schaik (Eds.), Cooperation in primates and humans: Mechanisms and evolution (pp. 107–119). Berlin: Springer.
- Mitani, J. C., & Watts, D. P. (2001). Why do chimpanzees hunt and share meat? Animal Behaviour, 61, 915–924.
- Neiworth, J. J., Johnson, E. T., Whillock, K., Greenberg, J., & Brown, V. (2009). Is a sense of inequity an ancestral primate trait? Testing social inequity in cotton top tamarins (Saguinus oedipus). Journal of Comparative Psychology, 123, 10–17.
- Nowak, M. A., & Sigmund, K. (2005). Evolution of indirect reciprocity. Nature, 437, 1291–1298.
- Ohtsuki, H., & Iwasa, Y. (2006). The leading eight: Social norms that can maintain cooperation by indirect reciprocity. Journal of Theoretical Biology, 239, 435–444.
- Pankner, A., Suomi, S. J., Visalberghi, E., & Ferrari, P. F. (2009). Capuchin monkeys display affiliation towards humans who imitate them. Science, 325, 880–883.
- Pierce, J., & Bekoff, M. (2012). Wild justice redux: What we know about social justice in animals and why it matters. Social Justice research, 25, 122–139.
- Povinelli, D. J. (1998). Can animals empathize? Maybe not. Scientific Americans (Feature article— Animal self-awareness: A debate). http://www.sciam.com/1998/1198intelligence/.
- Preston, S. D., & de Waal, F. B. M. (2002). Empathy: Its ultimate and proximate bases. Behavioral and Brain Sciences, 25, 1–72.
- Price, S., & Brosnan, S. F. (2012). To each according to his need? Variability in the responses to inequity in non-human primates. Social Justice Research, 25, 140–169.
- Raihani, N. J., Grutter, A. S., & Bshary, R. (2010). Punishers benefit from third-party punishment in fish. Science, 327, 171.
- Raihani, N. J., & McAuliffe, K. (2012). Does inequity aversion motivate punishment? Cleaner fish as a model system. Social Justice Research, 25, 213–231.
- Rand, D. G., Dreber, A., Ellingsen, T., Fudenberg, D., & Nowak, M. A. (2009). Positive interactions promote public cooperation. Science, 325, 1272–1276.
- Range, F., Leitner, K., & Vira´nyi, Z. (2012). The influence of the relationship and motivation on inequity aversion in dogs. Social Justice Research, 25, 170–194.
- Russell, Y. I., Call, J., & Dunbar, R. I. M. (2008). Image scoring in great apes. Behavioural Processes, 78, 108–111.
- Sanefuji, W., Ohgami, H., & Hashiya, K. (2006). Preference for peers in infancy. Infant Behavior and Development, 29, 584–593.
- Shimada, M. (2006). Social object play among young Japanese macaques (Macaca fuscata) in Arashiyama, Japan. Primates, 47, 342–349.
- Silberberg, A., Crescimbene, L., Addessi, E., Anderson, J. R., & Visalberghi, E. (2009). Does inequity aversion depend on a frustration effect? A test with capuchin monkeys (Cebus apella). Animal Cognition, 12, 505–509.
- Silk, J. B., Brosnan, S. F., Vonk, J., Henrich, J., Povinelli, D. J., Richardson, A. S., et al. (2005). Chimpanzees are indifferent to the welfare of unrelated group members. Nature, 27, 1357–1359.
- Singer, T. (2009). Understanding others: Brain mechanisms of theory of mind and empathy. In P. W. Glimcher, C. F. Camerer, E. Fehr, & R. A. Poldrack (Eds.), Neuroeconomics (pp. 251–268). New York: Academic Press.
- Singer, T., Seymour, B., O'Doherty, J. P., Stephan, K. E., Dolan, R. J., & Frith, C. D. (2006). Empathic neural responses are modulated by the perceived fairness of others. Nature, 439, 466–469.
- Skerry, A. E., Sheskin, M., & Santos, L. R. (2011). Capuchin monkeys are not prosocial in an instrumental helping task. Animal Cognition, 14, 647–654.
- Skitka, L. (2012). Cross-disciplinary conversations: A social psychological perspective on justice research with non-human animals. Social Justice Research, 25(3). doi:10.1007/s11211-012-0161-z.
- Stevens, J. R. (2010). Donor payoffs and other-regarding preferences in cotton-top tamarins (Saguinus oedipus). Animal Cognition, 13, 663–670.
- Subiaul, F., Vonk, J., Okamoto-Barth, S., & Barth, J. (2008). Do chimpanzees learn reputation by observation? Evidence from direct and indirect experience with generous and selfish strangers. Animal Cognition, 11, 611–623.
- Svetlova, M., Nichols, S. R., & Brownell, C. A. (2010). Toddlers' prosocial behavior: From instrumental to empathic to altruistic helping. Child Development, 81, 1814–1827.
- Takimoto, A., & Fujita, K. (2011). I acknowledge your help: Capuchin monkeys' sensitivity to others' labor. Animal Cognition, 14, 715–725.
- Takimoto, A., Kuroshima, H., & Fujita, K. (2010). Capuchin monkeys (Cebus apella) are sensitive to others' reward: An experimental analysis of food-choice for conspecifics. Animal Cognition, 13, 249–261.
- Talbot, C. F., Freeman, H. D., Williams, L. E., & Brosnan, S. F. (2011). Squirrel monkeys' response to inequitable outcomes indicates a behavioural convergence within the primates. Biology Letters, 7, 680–682.
- Thierry, B. (2000). Covariation of conflict management patterns across macaque species. In F. Aureli & F. B. M. de Waal (Eds.), Natural conflict resolution (pp. 106–128). Berkeley: University of California Press.
- Tomonaga, M., Tanaka, M., Matsuzawa, T., Myowa-Yamakoshi, M., Kosugi, D., Mizuno, Y., et al. (2004). Development of social cognition in infant chimpanzees (Pan troglodytes): Face recognition, smiling, gaze and the lack of triadic interactions. Japanese Psychological Research, 46, 227–235.
- Trivers, R. L. (1971). The evolution of reciprocal altruism. The Quarterly Review of Biology, 46, 25–57.
- Ueno, A., & Matsuzawa, T. (2004). Food transfer between chimpanzee mothers and their infants. Primates, 45, 231–239.
- van Leeuwen, E., Zimmermann, E., & Davila Ross, M. (2010). Responding to inequities: Gorillas try to maintain their competitive advantage during play fights. Biology Letters. doi:10.1098/rsbl.2010.0482.
- van Schaik, C. P., & van Hooff, J. A. R. A. M. (1996). Towards and understanding of the orangutan's social system. In W. C. McGrew, L. F. Marchant, & T. Nishida (Eds.), Great Ape Societies (pp.
- 3–15). Cambridge: Cambridge University Press.
- van Wolkenten, M., Brosnan, S. F., & de Waal, F. B. M. (2007). Inequity responses in monkeys are modified by effort. Proceedings of the National Academy of Sciences, USA, 104, 18854–18859.
- Vonk, J., Brosnan, S. F., Silk, J. B., Henrich, J., Richardson, A. S., Lambeth, S. P., et al. (2008). Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members. Animal Behaviour, 75, 1757–1770.
- Warneken, F., Hare, B., Melis, A. P., Hanus, D., & Tomasello, M. (2007). Spontaneous altruism by chimpanzees and young children. PLoS Biology, 5(7), e184. doi:10.1371/journal.pbio.0050184.
- Warneken, F., & Tomasello, M. (2006). Altruistic helping in human infants and young chimpanzees. Science, 311, 1301–1303.
- Yamagishi, T., Horita, Y., Takagishi, H., Shinada, M., Tanida, S., & Cook, K. S. (2009). The private rejection of unfair offers and emotional commitment. Proceedings of the National Academy of Sciences, USA, 106, 11520–11523.
- Yamamoto, S., Humle, T., & Tanaka, M. (2009). Chimpanzees help each other upon request. PLoS ONE, 4(10), e7416. doi:10.1371/journal.pone.0007416.
- Yamamoto, S., Humle, T., & Tanaka, M. (2012). Chimpanzees' flexible targeted helping based on an understanding of conspecifics' goals. Proceedings of the National Academy of Sciences, USA, 109, 3588–3592.
- Yamamoto, S., & Tanaka, M. (2009a). How did altruistic cooperation evolve in humans? Perspectives from experiments on chimpanzees (Pan troglodytes). Interaction Studies, 10, 150–182.
- Yamamoto, S., & Tanaka, M. (2009b). Do chimpanzees (Pan troglodytes) spontaneously take turns in a reciprocal cooperation task? Journal of Comparative Psychology, 123, 242–249.
- Yamamoto, S., & Tanaka, M. (2009c). Selfish strategies develop in social problem situations in chimpanzee (Pan troglodytes) mother–infant pairs. Animal Cognition, 12, S27–S36.
- Yamamoto, S., & Tanaka, M. (2010). The influence of kin relationship and a reciprocal context on chimpanzees' (Pan troglodytes) sensitivity to a partner's payoff. Animal Behaviour, 79, 595–602.
Figure 1

Two test paradigms for testing animals’ prosocial behavior. a Prosocial choice test animals were presented with two options: one is just self-rewarding, and the other benefits both the actor and the partner. In this picture, a subject (right) is choosing prosocial option (back) in which both the subject and the recipient received a high-value reward (photograph by Ayaka Takimoto). b Targeted helping test animals were tested whether they help others in predicament. In this picture, a chimpanzee (front) is giving a tool upon request to the partner who can thereafter use the tool to get otherwise out-of-reach reward. (photograph by Shinya Yamamoto)
Table 1

Figure 2
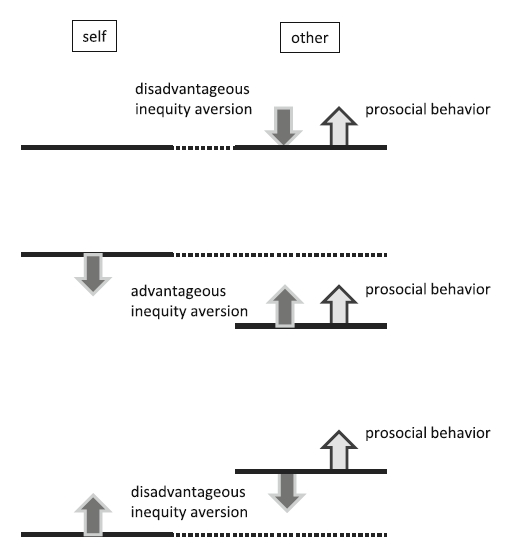
Response to equity and inequity of prosocial behavior and a sense of fairness. Top when there is equity between self and others, prosocial behavior generates disadvantageous inequity situation (light-gray arrow), and therefore, a sense of fairness may work the other way (dark-gray arrow). Middle when there is advantageous inequity situation, prosocial behavior and advantageous inequity aversion work to diminish the inequity. Bottom when there is disadvantageous inequity situation, prosocial behavior increases the inequity. Disadvantageous inequity aversion may restrict this unnecessarily excessive prosocial behavior
Figure 3
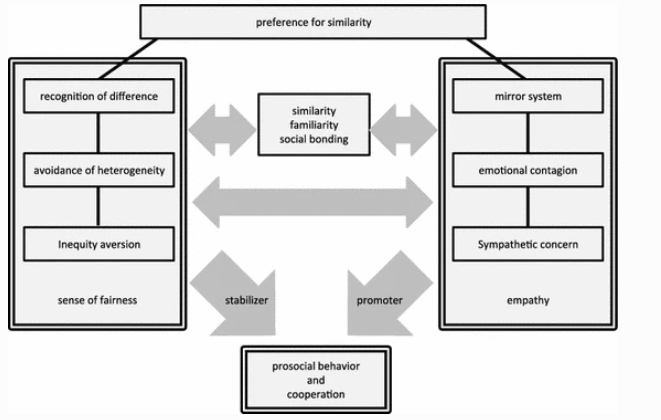
The relationship among prosocial behavior, a sense of fairness, empathy, and other relevant factors. A sense of fairness and empathy interact with each other (we do not mean that each factor in the two lines is linked one by one). Similarity also influences these two mechanisms, and therefore ultimately has an effect on prosocial behavior