The influence of kin relationship and reciprocal context on chimpanzees' other-regarding preferences.
Shinya Yamamoto, Masayuki Tanaka
DOI: 10.1016/j.anbehav.2009.11.034Abstract
We investigated the evolutionary origin of other-regarding preferences, one of the strong underlying motivations for altruism, in the chimpanzee, Pan troglodytes. Although altruism is expected theoretically to be kin biased and frequent in a reciprocal context, few experimental studies to date have specifically tested these hypotheses from the viewpoint of proximate mechanisms. We examined the other-regarding preferences of individuals in mother–offspring pairs and in nonkin adult pairs in both reciprocal and nonreciprocal contexts. Based on the previously established choice paradigm with mutually or selfishly beneficial options, we developed a novel task using buttons. In experiment 1, chimpanzee participants involving three mother–offspring pairs were offered two options: delivering food rewards to their partner and themselves or only to themselves. We compared their choices between partner-present and partner-absent conditions. In experiment 2, we developed a reciprocal context in which the two participants alternately chose the two options. In contrast to the theoretical predictions, the chimpanzees did not show any prosocial tendencies even between mother and offspring or in a reciprocal context. We propose that the experimental set-up which prevented direct interactions between the participants might have influenced these results. In conclusion, the present study suggests that voluntary and/or strategic other-rewarding behaviour arose in humans after divergence from the common ancestor of humans and chimpanzees.
Keywords
altruism, chimpanzee, mother-offspring relationship, nepotism, other-regarding preference, Pan troglodytes, prosociality, proximate mechanism, reciprocity,
We investigated the evolutionary origin of altruism in chimpanzees, Pan troglodytes, one of our closest living relatives, from the viewpoint of proximate mechanisms. Previously, the evolution of altruism has been explained mainly by the theories of kin selection (Hamilton 1964) and reciprocal altruism (Trivers 1971). However, these kinds of theoretical framework normally deal with the ultimate reasons, providing an explanation for the fitness benefits derived from altruism, and fail to explain how such prosociality is achieved. The proximate mechanism and its evolutionary origins still remain to be examined.
With this proximate perspective, recent comparative cognitive studies with various primates have examined whether nonhuman animals are sensitive to others' payoffs. Research has especially focused on other-regarding preferences, which underlie motivation for altruism in humans. Common marmosets, Callithrix jacchus (Burkart et al. 2007) and capuchin monkeys, Cebus apella ( [Lakshminarayanan and Santos, 2008], [de Waal et al., 2008] and [Takimoto et al., In press]) clearly show other-regarding preferences, that is, spontaneously sharing food with nonreciprocating and genetically unrelated individuals.
However, controversy still abounds as to whether or not chimpanzees also show such other-regarding preferences. Both observational studies ( [Goodall, 1986] and [de Waal, 1996]) and experimental studies ( [Warneken and Tomasello, 2006], [Warneken et al., 2007] and [Yamamoto et al., 2009]) have yielded evidence for prosocial and cooperative behaviour in chimpanzees. Meanwhile, other experimental studies have failed to show they have other-regarding preferences. When chimpanzees were offered a choice between two options, a mutually beneficial option or a selfishly beneficial option, they did not change their choice whether a conspecific partner was present or absent ( [Silk et al., 2005], [Jensen et al., 2006] and [Vonk et al., 2008]). In a review of these studies, Yamamoto & Tanaka (2009a) suggested that chimpanzees' prosocial behaviour would not occur voluntarily but required explicit recipient's request or solicitation (see also supporting experiments: [Warneken and Tomasello, 2006], [Warneken et al., 2007] and [Yamamoto et al., 2009]).
Now the question is, do chimpanzees show other-regarding preferences if unable to interact, or are prevented from interacting, with one another? In the present study, we examined chimpanzees' other-regarding preferences in two contexts theoretically proposed to favour the evolution of altruism: between kin individuals and in a context of reciprocity. Previous studies normally tested nonkin pairs in a unilateral condition in which participants could not expect any reciprocation from their partner. However, considering that altruism is theoretically predicted to have evolved among kin and/or through a system of reciprocity, other-regarding preferences might be more apparent under such conditions.
Regarding nepotism in chimpanzees, helping behaviour seems to be biased from mothers towards their offspring ( [Goodall, 1986] and [Hirata, 2009]). However, some studies have reported limited impact of kinship on cooperative relationships in wild chimpanzees ( [Mitani et al., 2000] and [Langergraber et al., 2007]) and on other-regarding preferences in captive chimpanzees (Jensen et al. 2006). In addition, a detailed experimental analysis of food sharing between mothers and infant chimpanzees showed that even mothers rarely transfer edible parts voluntarily to their 1-year-old infant (Ueno & Matsuzawa 2004). It still remains to be examined whether chimpanzee mothers are sensitive to their offspring's payoff and vice versa. To examine this, we tested mother–offspring chimpanzee pairs (in which the offspring were raised by their biological mothers) as well as nonkin adult pairs.
Reciprocity is also observed in chimpanzees; however, the underlying motivations are still unclear. Observational studies have suggested reciprocal correlations in giving and receiving commodities and services, such as food, grooming, agonistic support and mating ( [de Waal, 1997] and [Mitani and Watts, 2001]; Koyama et al. 2006; Duffy et al. 2007; Hockings et al. 2007; but see also criticisms: [Hammerstein, 2003], [Stevens, 2004], [Stevens and Gilby, 2004] and [Gilby, 2006]). However, previous experimental studies with controlled settings have mostly failed to confirm chimpanzees' reciprocal cooperation. For example, chimpanzees failed to take turns spontaneously in performing other-rewarding behaviour ( [Hikami and Matsuzawa, 1991], [Yamamoto and Tanaka, 2009b] and [Yamamoto and Tanaka, 2009c]), to trade valuable tokens ( [Brosnan and Beran, 2009] and [Pele et al., 2009]) or to reciprocate a partner's prosocial behaviour (Brosnan et al. 2009). Although Melis et al. (2008) found that chimpanzees in a mutually beneficial cooperation task chose a partner who had previously behaved favourably towards them, the evidence for reciprocity was very weak, and again we cannot tell whether chimpanzees' other-regarding sentiments prompted their choice. At this point, we know little about how reciprocity in behaviour emerges between chimpanzees. Hence it would be valuable to examine chimpanzees' other-regarding preferences in reciprocal contexts.
We tested chimpanzees' other-regarding preferences in a newly developed task, modifying the previously devised paradigm with 1/0 (selfish) and 1/1 (prosocial) choices ( [Silk et al., 2005], [Jensen et al., 2006] and [Vonk et al., 2008]). In this experimental paradigm, a participant could give a food reward to a conspecific partner by choosing a 1/1 option without any cost. This led to the expectation that if a participant was positively other-regarding, the participant would choose the 1/1 option more when a partner was present than when a partner was absent. In contrast, if a participant was aversive to inequity or spiteful, it would choose the 1/0 option more often in a partner-present condition. Previous studies ( [Silk et al., 2005], [Jensen et al., 2006] and [Vonk et al., 2008]) adopted intuitive tasks; for example, food rewards were presented on trays in front of the participants, and they pulled a bar which was connected to one of the food trays. However, chimpanzees have more difficulties in inhibitory control when faced with such intuitive tasks than with equivalent cognitive tasks ( [Boysen and Berntson, 1995], [Boysen et al., 1996] and [Boysen et al., 1999]). In previous studies, the chimpanzees might have focused their attention on their own reward but not on the partner's. Therefore, in the present study, to overcome inhibitory control problems, we developed a more cognitively demanding task using buttons whose association with food rewards was less intuitive. We developed an apparatus and procedures that drew the participant's attention to his/her partner's payoff.
Experiment 1
We investigated chimpanzees' other-regarding preferences using a novel paradigm and task. Experiment 1 was designed to examine the influence of nepotism, especially in mother–offspring pairs, on chimpanzees' other-regarding preferences.
Methods
Participants
Participants were socially housed chimpanzees at the Primate Research Institute (PRI), Kyoto University, Japan. All participants had taken part in a variety of tests on perception and cognition ( [Matsuzawa, 2003], [Matsuzawa, 2009] and [Matsuzawa et al., 2006]), including social tasks involving contexts of food sharing (Ueno & Matsuzawa 2004), token sharing (Tanaka & Yamamoto 2009) and reciprocity ( [Hikami and Matsuzawa, 1991], [Yamamoto and Tanaka, 2009b] and [Yamamoto and Tanaka, 2009c]); but the test paradigm for the present study was novel to the participants, and this study was conducted at least 4 months after the previous social tasks. The participants spent their daily life with other group members. The group consisted of 13 chimpanzees (three males and 10 females) of three generations. The housing facilities comprised nine indoor rooms and three outdoor compounds (approximately 700 m2, 100 m2 and 100 m2) with several climbing structures (a maximum of 15 m), small streams and abundant vegetation (Ochiai & Matsuzawa 1998). The chimpanzees had ad libitum access to water and were not food deprived during the study. They were given monkey pellets and various kinds of seasonal fruits and vegetables at least three times a day (breakfast, lunch and dinner), and additionally a maximum of four times during experiments. The chimpanzees were tested and cared for according to PRI's own guidelines for the ethical conduct of research.
Ten chimpanzees including three mother–offspring pairs participated in the present study, although one adult male was later excluded from the analysis because he failed to demonstrate an understanding of the apparatus. The three offspring were born in 2000 at PRI, and were raised by their biological mothers. The offspring were 6 years old at the start of the study, and still somewhat dependent on their mother; for example, they always came to the experimental room with their mother and refused to come alone. Each participant was paired with his/her mother (N = 3), her offspring (N = 3) or a nonkin partner (N = 3), and played both the roles of actor and recipient (for details, see Table 1). We chose the above 10 participants and paired combinations for the test, based on the chimpanzees' motivation to participate in our study and their personal compatibility; for example, we did not form pairs of males, in case they became aroused in the test booths.
| Actor | Sex/age (years) | Paired with | Kinship | r |
|---|---|---|---|---|
| Ayumu | Male/6 | Ai | Filial | 0.5 |
| Cleo | Female/6 | Chloe | Filial | 0.5 |
| Pal | Female/6 | Pan | Filial | 0.5 |
| Ai | Female/30 | Ayumu | Maternal | 0.5 |
| Chloe | Female/26 | Cleo | Maternal | 0.5 |
| Pan | Female/23 | Pal | Maternal | 0.5 |
| Mari | Female/30 | Akira | None | 0 |
| Akira | Male/30 | Penedesa | None | 0 |
| Pendesa | Female/30 | Gon | None | 0 |
| Gon | Male/40 | Mari, but excluded from the analysis | ||
r: degree of genetic relatedness.
Apparatus and set-up
We developed an apparatus and set-up for the selfish versus prosocial choice task using buttons (Fig. 1). The chimpanzee participants were tested in an experimental room with three adjacent booths (actor's booth: 136 cm × 142 cm; recipient's booth: 291 cm × 192 cm; control booth: 155 cm × 142 cm). The walls and partitions consisted of transparent polycarbonate panels. The distance between food boxes in the actor's and recipient's booths was approximately 1 m. An actor could see food delivery not only to his or her own booth but also to that of the recipient.

Figure 1. The experimental set-up. In this picture, the chimpanzee in the foreground booth (the actor's booth) is offered a choice of buttons between two options: the 1/0 option (left orange button) and the 1/1+ option (right red button). The partner chimpanzee in the background booth (the recipient's booth) is a potential recipient. The control booth is not shown here, but was located behind the actor's booth (on the left outside of the picture) separated by transparent panels.
In the actor's booth, there were three illuminated push-button switches (Sunmulon Co., Ltd., Tokyo, Japan; illuminated switch XH) on a wall: one button was the start key, and the other two were for the choice stimulus. The start key button flashed blue, and the choice buttons flashed in one of three colours: green, red or orange. Each button was equipped with three flash bulbs of different colour, so we could randomize the position of the choice buttons across trials. A button that flashed green (green button) meant a 0/0 option which delivered no reward to either booth. This 0/0 option was incorporated to prevent the actor from being indifferent to the colour of the choice button. Buttons that flashed red and orange (red button and orange button) were assigned either to the selfish option which delivered a piece of apple (1 cm cube) only to the actor's booth (1/0 option, hereafter), or to the prosocial option which delivered a piece of apple to the actor's booth as well as a piece of apple and a quarter of a grape to the recipient's booth (1/1+ option; see also Procedure below for more details). We used grapes, which the chimpanzees preferred to apple, as a reward to a recipient, to call the actor's attention to the recipient's payoff.
The food rewards were delivered via universal feeders (Biomedica Ltd., Osaka, Japan BUF310-P50) with a concurrent chime indicating food delivery and its location (different chimes for the actor's booth and the recipient's booth). Reward to the recipient's booth was delivered just after the actor's choice of the 1/1+ option, while reward to the actor's booth was always delivered 2 s after the actor's choice of the 1/1+ or 1/0 option. We set this time lag to make the food delivery to the recipient's room more salient to the actor. The control booth was irrelevant to any reward. When a partner was in the control booth and when an actor chose the 1/1+ option, pieces of apple and grape were delivered to the empty recipient's booth, although the food did not remain there but dropped out of the booth through a food box. A computer (Toshiba, PS426) running a Visual Basic 6.0 program controlled the buttons, the feeders and the chimes.
Procedure
The participants were tested in the actor's booth. Experiment 1 consisted of a training phase, a pretest knowledge phase (one session), a test phase (20 sessions for each of two conditions) and a post-test knowledge phase (one session). One session consisted of 54 trials (1/1+ versus 1/0: 12 trials; 1/1+ versus 0/0: 12 trials; 1/0 versus 0/0: 12 trials; 1/1+ versus 1/1+: 6 trials; 1/0 versus 1/0: 6 trials; 0/0 versus 0/0: 6 trials; the order was random and the left–right position was counterbalanced in each session), although the main target of the analysis was choices in trials 1/1+ versus 1/0. Each trial started when the start button flashed with a concurrent chime (‘Pi’). Just after the actor pushed the start button, the two choice buttons flashed. A trial ended when the actor pushed either choice button or after 5 s had passed since the choice buttons flashed. The intertrial interval was 5 s. All the procedures during a session were controlled by a computer program without any intervention of human experimenters.
In the training phase, the participants were individually trained to understand the set-up and distribution of rewards for each coloured button. In this phase, there was no partner, and the partition between the actor's booth and the recipient's booth was open, so that the participants could move freely between the two booths. Therefore we expected the participants to choose the 1/1+ option rather than the 1/0 option if they understood that the 1/1+ choice delivered rewards both to the actor's booth and to the recipient's booth while the 1/0 choice generated reward only to the actor's booth. The criteria for passing this training phase was that a participant chose the 1/1+ option significantly more often than the 1/0 option in 1/1+ versus 1/0 trials (10 or more out of 12 trials; binomial test: P < 0.05) during three or more consecutive sessions.
After passing the training phase, on another day, we confirmed the participants' knowledge in the same situation as the training phase but with a partner in the control booth (pretest knowledge phase). We examined whether the participants' performance was maintained in this situation with a partner present, and judged the participants' knowledge as adequate if they chose the 1/1+ option significantly more often than the 1/0 option in 1/1+ versus 1/0 trials (in 10 or more of 12 trials). In the training phase and the pretest knowledge phase, a red button was assigned as the 1/1+ option and an orange button was assigned as the 1/0 option.
In the test phase, we created two conditions: partner-present and partner-absent. In both conditions, a partner was present, but the partner was in the recipient's booth in the partner-present condition and in the control booth in the partner-absent condition. Both conditions were conducted for one session each for each pair once a day. The order of the conditions was counterbalanced across sessions. The red button was 1/1+ and the orange button was 1/0 during the first 10 sessions for each condition, and then the colours were reversed in the second set of 10 sessions for each condition: the red button became the 1/0 option and the orange one the 1/1+ option. We adopted this switching procedure between the first and second halves in case the tenacity of a trained choice (red button) could explain the participants' preference for the 1/1+ option and hide differences between the two conditions in the test phase.
After the test phase, we checked the participants' understanding of the reversed distribution of rewards for red- and orange-coloured options (post-test knowledge phase). This phase was identical to the pretest knowledge phase except that the red button was 1/0 and the orange button was 1/1+ in this post-test knowledge phase.
To prevent any possibility for immediate reciprocity, tests for the mothers and those for their offspring were separated temporally; first the offspring was the actor and the mother was the potential recipient, and then after more than 1 week, the roles were reversed. This kind of problem with reciprocation did not arise among the nonkin adult pairs, since each participant was paired with a different partner when he or she was an actor or a recipient.
Coding and analysis
The participants' choice was automatically recorded by the computer controlling the apparatus. The data were analysed at both the group and individual level. To conduct the group level analysis, we compared the participants' choices between the two conditions and between the two phases. To determine whether any individual chimpanzee showed a difference in their choice between the partner-present and partner-absent conditions, we performed a Wilcoxon signed-ranks test on the data for each participant. Each session of the partner-present and partner-absent conditions on a same day was paired, and the sample size was 10 (the number of sessions of each condition) for each participant. We also compared the performance between kin pairs and nonkin pairs. We used nonparametric tests throughout, since we could not assume a normal distribution owing to the small sample size. All analyses were done using SPSS 13.0 (SPSS Inc., Chicago, IL, U.S.A.). All tests were two tailed, and the level of statistical significance was set at 0.05. All values reported are mean ± SE.
Results
In the training phase, all the participants except one male (Gon) met the criteria in an average of 6.1 ± 0.73 sessions. Gon failed to meet the criteria even after 40 sessions. We excluded his data in test and knowledge phases from the analysis, although he participated in all the phases as a partner to Mari and Pendesa (see Table 1). By the time the participants met the performance criteria, all the participants avoided the 0/0 option when it was presented alongside the 1/1+ or 1/0 option. We confirmed the participants' understanding on another day (the pretest knowledge phase) just before the test phase, and all the participants met the criteria; that is, they preferred the 1/1+ option (red button) to the 1/0 option (orange button) in an average of 94.4 ± 2.8% of their choices.
Overall, in the test phase, there was no significant difference in the actor's preference for the 1/1+ option over the 1/0 option between partner-present and partner-absent conditions (Fig. 2). In the first 10 sessions, we predicted that the participants would decrease their 1/1+ choice (red button) only in the partner-present condition if they were aversive to inequity or spiteful. Nevertheless, in this situation, participants maintained a high preference for the 1/1+ option. In addition there was no significant difference between the partner-present and partner-absent conditions (partner-present: 95.1 ± 1.9%; partner-absent: 94.6 ± 2.4%; Wilcoxon signed-ranks test: Z = −0.67, P = 0.63). There was also no significant difference between the pretest knowledge phase and the first 10 sessions of the test phase (the partner-absent condition; Wilcoxon signed-ranks test: Z = −0.42, P = 0.71). In the second set of 10 sessions after reversing the function of the red and orange buttons (thereafter red: 1/0, orange 1/1+), we predicted that the participants would increase their 1/1+ choice (orange button) only in the partner-present condition if they exhibited positive other-regarding preferences. There was no significant difference between the two conditions (partner-present: 11.0 ± 4.7%; partner-absent: 10.1 ± 4.7%; Wilcoxon signed-ranks test: Z = −1.36, P = 0.22).
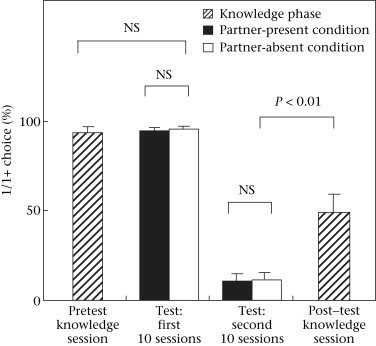
Figure 2. Percentage of trials in experiment 1 in which chimpanzee actors chose the 1/1+ option over the 1/0 option. A red button was assigned as the 1/1+ option and an orange button as the 1/0 option in the pretest knowledge session and the first 10 sessions in the test phase. Thereafter the colours were reversed: a red button was assigned as the 1/0 option and an orange button as the 1/1+ option in the second 10 sessions in the test phase and in the post-test knowledge session. The bars represent SE, and Wilcoxon signed-ranks tests were used for the comparisons. N = 9 in each phase.
Individual analyses of the choice between the partner-present and partner-absent conditions were not significant for all participants (Wilcoxon signed-ranks test: all P values >0.25). This means that even chimpanzee mothers and offspring did not show any significant difference in their choice between the two conditions. We also compared the ratios of 1/1+ choice in the partner-present condition between when paired with a kin partner (mother or offspring) and when paired with a nonkin partner, and found no significant difference (the first 10 sessions: when paired with a kin partner (N = 6): 94.8 ± 2.6%; when paired with a nonkin partner (N = 3): 95.8 ± 3.4%; Mann–Whitney U test: U = 8.5, P = 0.89; the second 10 sessions: when paired with a kin partner (N = 6): 6.4 ± 4.3%; when paired with a nonkin partner (N = 3): 20.4 ± 10.3%; Mann–Whitney U test: U = 5, P = 0.29).
In the post-test knowledge phase (just one session), all the participants except one female (Chloe) increased the ratio of 1/1+ choice compared to that in the test phase (the second 10 sessions). There was a significant difference between the two (the last 10 sessions of the test phase (the partner-absent condition): 10.1 ± 4.7%; the post-test knowledge phase: 49.1 ± 10.8%; Wilcoxon signed-ranks test: Z = −2.5, P = 0.008). Five of the nine participants chose the 1/1+ option on the first trial of the post-test knowledge phase. We compared the participants' 1/1+ choice between the first six trials and the second six trials to examine the effect of learning during the post-test knowledge phase. This analysis showed no significant difference in performance between the two (the first six trials: 48.1 ± 12.6%; the second six trials: 50.0 ± 9.6%; Wilcoxon signed-ranks test: Z = −0.11, P = 1.0).
Discussion
These results suggest that chimpanzees do not show any other-regarding preferences in this paradigm using 1/1+ and 1/0 choices, which concords with the conclusions of previous studies ( [Silk et al., 2005], [Jensen et al., 2006] and [Vonk et al., 2008]). Experiment 1 showed that there was no difference between the partner-present and partner-absent conditions. Although the participants showed overall preferences for the red button (1/1+ in the first 10 sessions and 1/0 in the second 10 sessions), this can be interpreted as persistence from the training, since the red button was assigned to the 1/1+ option in the training phase. In the partner-present condition compared to the partner-absent condition, we found no decrease in 1/1+ choice in the first 10 sessions and no increase in the second 10 sessions after switching the functions of the red and orange buttons. These results suggest that the chimpanzees showed neither inequity aversion nor positive other-regarding preferences. Considering previous findings concerning chimpanzees' sense of fairness (Brosnan et al. 2005; but also see [Brauer et al., 2006] and [Jensen et al., 2007]), it might be possible that the participants showed both inequity aversion and positive other-regarding preferences, and that these two tendencies were offset. However, it seems unlikely that inequity aversion and positive other-regarding preferences were absolutely balanced and offset in all the nine participants.
The results of experiment 1 indicate the absence of other-regarding preferences even between mothers and offspring. This conclusion apparently contradicts naturalistic observations that chimpanzee mothers behave altruistically towards their offspring (e.g. [Goodall, 1986] and [Hirata, 2009]). However, detailed analysis of mother–offspring interactions have shown that chimpanzee mothers' attitude towards their offspring can be characterized as tolerance rather than active altruism ( [Matsuzawa et al., 2001], [Ueno and Matsuzawa, 2004] and [Yamamoto and Tanaka, 2009c]). At the very least, the results from the kin pairs in the present study suggest that chimpanzees, even mothers and offspring, do not show unconditional other-regarding preference.
The performance in the two knowledge phases, especially the post-test knowledge phase, indicated that the participants (except Chloe) understood the apparatus and were able to distinguish between the 1/1+ and 1/0 options. In only one session of the post-test knowledge phase did the participants significantly increase their 1/1+ choice compared with during the previous test phase, although there was still some persistence in the red button (the 1/0 option). The results also show that the participants did not learn the button functions during the post-test knowledge phase. It seems that the participants had understood the button functions by the test phase even after the functions of the red and orange buttons were switched. The participants' avoidance of the green button (the 0/0 option) also supports the idea that the participants paid attention to the colour of the buttons.
Experiment 2
We conducted experiment 2 at least 1 month after experiment 1. The participants and the pairings were the same as in experiment 1 (see Table 1). One male (Gon) was again eliminated from the analysis, although he participated in the test.
The apparatus and set-up were slightly modified from experiment 1. We devised a set of three buttons (one start key button and two choice buttons) not only on a wall in the actor's booth but also on a wall in the recipient's booth. We offered to the participants the two options, 1/1+ and 1/0, and excluded the 0/0 option. An orange button was assigned as the 1/1+ option and a red button was assigned as the 1/0 option throughout experiment 2. The participants could see the partner pushing a button and receiving food rewards through the transparent panels.
Experiment 2 consisted of a test phase and a post-test knowledge phase. We did not carry out any additional training and pretest knowledge phases before experiment 2 to avoid biasing the chimpanzees to choose the 1/1+ option. The unilateral condition was similar to the partner-present condition in experiment 1: only the actor had the choice of the 1/1+ or 1/0 options throughout a session. In contrast, in the bilateral condition, both the actor and the recipient made a choice on alternate trials. A session started with the recipient's choice, and then the actor made a choice and vice versa. A session consisted of 36 trials in the unilateral condition and 72 trials (36 choices for each participant) in the bilateral conditions, including 12 and 24 warm-up trials, respectively, at the beginning. In the warm-up trials, each participant (or only an actor in the unilateral condition) was offered six 1/1+ versus 1/1+ choices (trials) and six 1/0 versus 1/0 choices, so that the participants had opportunities to choose each option, to understand the reward distribution of each option. The participants in the bilateral condition also had experience of the partner's choice of 1/1+ and 1/0 during warm-up trials. One session with the unilateral condition and one session with the bilateral condition were conducted for each pair in a day, and the order of the two conditions was counterbalanced across sessions. We conducted 10 sessions for each condition. We did not conduct a nonsocial control in experiment 2, since our aim was to investigate the influence of a reciprocal context upon chimpanzees' other-regarding preferences.
In the post-test knowledge phase, we checked the participants' understanding of the apparatus and of the reward distribution of the options. Each participant was offered one session of 24 1/1+ versus 1/0 choices with the partition open between the actor's and the recipient's booths and with a partner in the control booth. In both phases, the right–left position of the 1/1+ and 1/0 options was counterbalanced within a session.
We focused on the actors' choice for the analysis. The data were analysed at both the group and individual level. We first investigated whether or not there was any difference in the actors' choice between the two conditions (the unilateral and bilateral conditions) and between the two phases (the test phase and the post-test knowledge phase). Then we tested whether or not an actor's choice in the bilateral condition was influenced by the preceding partner's (recipient's) choice. We also compared the performance between kin pairs and nonkin pairs as in experiment 1.
Results
Contrary to our prediction, we found no significant difference in the actors' choice between the unilateral and bilateral conditions in the test phase (Fig. 3a). The participants chose the 1/1+ option in, on average, 64.8 ± 7.5% of trials in the unilateral condition and 64.3 ± 6.6% in the bilateral condition. There was no significant difference between the two conditions (Wilcoxon signed-ranks test: Z = −0.059, P = 1.0). This result was true for all the participants, regardless of the partner, whether the mother, the offspring or a nonkin individual (Wilcoxon signed-ranks test: all P values >0.05).
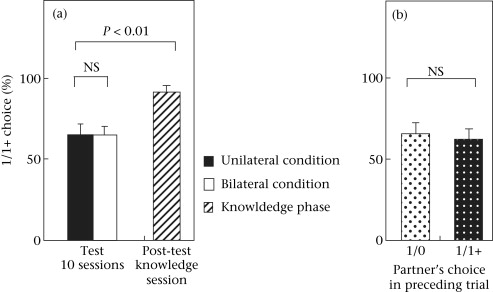
Figure 3. Percentage of trials in experiment 2 in which chimpanzee actors chose the 1/1+ option over the 1/0 option. The bars represent SE, and Wilcoxon signed-ranks tests were used for the comparisons. (a) Examination of the influence of reciprocal (bilateral condition) and nonreciprocal (unilateral condition) contexts over actors' choice in the test phase and in comparison with the knowledge phase. N = 9 in each phase. (b) Examination of the influence of the preceding partner's choice (1/0 or 1/1+) over the actors' choice. N = 9 for each choice option.
We also found no influence of the preceding partner's choice on the actors' choice in the bilateral condition (Fig. 3b). After the partner chose the 1/0 option, the participants in the next trial chose the 1/1+ option on average 66.3 ± 7.1% of the time. After the partner chose the 1/1+ option, the participants in the next trial chose the 1/1+ option on average 62.7 ± 6.8% of the time. There was no significant difference between these two values (Wilcoxon signed-ranks test: Z = −0.53, P = 0.65). All the participants, whether paired with their mother, their offspring or a nonkin individual, showed no significant change in their choice according to the preceding partner's choice (Fisher's exact test: all P values >0.26). We compared the ratios of 1/1+ choice between when paired with a kin partner (mother or offspring) and when paired with a nonkin partner, and found no significant difference (the unilateral condition: when paired with a kin partner (N = 6): 62.4 ± 10.6%; when paired with a nonkin partner (N = 3): 69.6 ± 9.8%; Mann–Whitney U test: U = 8, P = 0.80; the bilateral condition: when paired with a kin partner (N = 6): 65.1 ± 9.3%; when paired with a nonkin partner (N = 3): 62.6 ± 9.2%; Mann–Whitney U test: U = 7, P = 0.61).
In the post-test knowledge phase (one session), all the participants increased the ratio of 1/1+ choices compared to that in the test phase (the unilateral condition). There was a significant difference between the two (the test phase (the unilateral condition): 64.8 ± 7.5%; the post-test knowledge phase: 91.7 ± 4.4%; Wilcoxon signed-ranks test: Z = −2.67, P = 0.004). Eight of the nine participants chose the 1/1+ option on the first trial of the post-test knowledge phase. We compared the participants' 1/1+ choice between the first 12 trials and the second 12 trials to examine the effect of learning during the post-test knowledge phase, and found no significant difference in performance between the two (the first 12 trials: 90.7 ± 5.1%; the second 12 trials: 91.7 ± 3.7%; Wilcoxon signed-ranks test: Z = −0.45, P = 1.0). In particular, six of the nine participants chose the 1/1+ option in all 24 trials in the post-test knowledge phase.
Discussion
Overall, the chimpanzees did not show any increase of their other-regarding preferences in the reciprocal context. In the bilateral condition, paired participants could have increased their own gain if they had chosen the 1/1+ option for each other. This kind of reciprocal cooperation was particularly needed for the participants to get the preferred reward (grape) which was delivered only when the partner chose the 1/1+ option in the bilateral condition. Nevertheless, the chimpanzees did not increase their cooperative choice favouring 1/1+ in the bilateral condition. This suggests that chimpanzees do not show strategic other-rewarding behaviour in reciprocal contexts. It might be difficult for chimpanzees to deduce that giving rewards to their partner would increase their own chances of receiving rewards thereafter.
We also found no influence of the preceding partner's choice over the chimpanzees' other-regarding preferences, which supports previous findings (Brosnan et al. 2009). Although previous studies suggested that cottontop tamarins, Saguinus oedipus (Hauser et al. 2003) and capuchin monkeys ( [de Waal, 2000] and [de Waal and Berger, 2000]) have some reciprocal inclination, and an even more sophisticated strategy such as two tits for a tat (in cottontop tamarins: Chen & Hauser 2005), we found no such evidence in our chimpanzee subjects. The results of experiment 2 suggest no reciprocal prosociality (if not altruism) in the chimpanzees.
The increased performance in favour of the 1/1+ option in the post-knowledge phase showed that the participants understood the apparatus and were able to distinguish between the two options. The results also suggest that the participants did not learn the button functions during this phase, but already understood them during the test phase. The turn taking in making a choice and the food delivery were salient to both participants both visually and acoustically (owing to the chimes). They could also watch their partner consume food rewards and pushing a button, since the partner sat only 1 m away across the transparent partition. In addition, all the participants had experience of choosing in both the actor's and recipient's booths. It is therefore unlikely that the participants failed to understand their partner's role as well as their own. The performance in the second condition might have been influenced by that in the first, since we conducted the two conditions on the same day. Nevertheless, the considerable number of test and warm-up trials ensured that the participants understood the difference between the two conditions.
General discussion
The present experimental study failed to reveal any significant influence of kin relationship and reciprocity on chimpanzees' other-regarding preferences. In our experiments using the 1/1+ versus 1/0 paradigm, the chimpanzees showed no other-regarding preferences whether between mother and offspring or in a reciprocal context. This suggests that chimpanzees, even kin, do not voluntarily share food with others either prosocially or strategically to elicit reciprocation.
The experimental set-up in the present study differs considerably from previous studies in which, for example, chimpanzees pulled a bar connected to a food tray. Nevertheless, the difference in set-up did not affect the chimpanzees' other-regarding preferences. By using a computer-controlled apparatus, we could eliminate any intervention by human experimenters and carry out a considerable number of trials (240 test trials per individual for each experiment). The payoff to a recipient was salient for the actor owing to the chime associated with food delivery and the time delay (food for a recipient was delivered 2 s before food delivery to an actor). The participant could also see food rewards drop from the feeder through transparent tubes to both the recipient's booth and his/her own. This cognitively demanding button choice task was designed to help overcome issues with inhibitory control (see [Boysen and Berntson, 1995], [Boysen et al., 1996] and [Boysen et al., 1999]). Even with these experimental treatments, we obtained no evidence of chimpanzees' voluntary prosociality.
The negative results might have come from the apparently unnatural set-up for chimpanzees. However, we confirmed the participants' understanding of the set-up in the knowledge phases. In addition, all the participants were already used to participating in experiments with artificial set-ups with monitors and vending machines ( [Matsuzawa, 2003], [Matsuzawa, 2009] and [Matsuzawa et al., 2006]). In fact, in our previous experiments using vending machines, which might have been intuitively complicated for chimpanzees, the participants understood the set-up and showed various interactions between them ( [Yamamoto and Tanaka, 2009b] and [Yamamoto and Tanaka, 2009c]). It is still possible that the chimpanzees did not necessarily understand how their choices influenced the partner. However, they clearly understood the functions of the 1/1+ and 1/0 options, and were able to adjust their choice to maximize their reward when they had free access to both the actor's and the recipient's booths. This result actually suggests that chimpanzees are more self-regarding than other-regarding.
The present study suggests the importance of perspectives on proximate mechanisms. Although our results contradict theoretical expectations, that is, kin-biased altruism and reciprocal altruism, they support the hypothesis that chimpanzees are seldom voluntarily altruistic especially in contexts involving food (Yamamoto & Tanaka 2009a). Previous studies have pointed out the importance of the recipient's request for altruism in chimpanzees ( [Warneken and Tomasello, 2006], [Warneken et al., 2007] and [Yamamoto et al., 2009]; reviewed in Yamamoto & Tanaka 2009a). If other-rewarding behaviour in chimpanzees occurs upon a recipient's request rather than voluntarily, this can explain the negative results obtained in the present study. The participants in the present study were separated by transparent partitions. Although they could have communicated with each other by making eye contact, they did not have any way to make their request explicitly. This study supports the idea that chimpanzees are not positively other-regarding without a recipient's request ( [Yamamoto and Tanaka, 2009a] and [Yamamoto et al., 2009]). Previous studies have also suggested the importance of a request even in mother–infant pairs (Ueno & Matsuzawa 2004) and in a reciprocal context (Yamamoto & Tanaka 2009b).
In sum, this study suggests that voluntary and/or strategic other-rewarding behaviour, especially food donation, evolved in humans after divergence from the common ancestor of humans and chimpanzees. Some evidence for prosociality has been reported in some catarrhines (common marmosets: Burkart et al. 2007; tufted capuchin monkeys: [de Waal et al., 2008] and [Lakshminarayanan and Santos, 2008], but not in cottontop tamarins: Cronin et al. 2009); however, the continuity between humans and these other primate taxa needs to be discussed carefully. It is highly possible that other-regarding preferences might have evolved in phylogenetically diverse taxa, although there seems to be no consensus at this point on what the decisive factors in explaining species differences are. At the very least, it is obvious from the present and previous studies ( [Silk et al., 2005], [Jensen et al., 2006] and [Vonk et al., 2008]) that chimpanzees do not show any prosocial tendency in experiments using the 1/1 (or 1/1+) and 1/0 paradigm with food without direct interaction between individuals. In future studies with chimpanzees, detailed analyses are necessary to examine more precisely under which conditions chimpanzees show prosociality and what kind of interactions between individuals actually elicit and maintain other-rewarding behaviour.
Acknowledgments
The present study was financially supported by a grant (no. 16002001 and no. 20002001 to T. Matsuzawa) from the Ministry of Education, Culture, Sports and Technology (MEXT), Japan, by the Global COE Program A06 and D07 to Kyoto University of MEXT, and by a grant from the Japan Society for the Promotion of Science for Young Scientists (no. 18-3451 and no. 21-9340 to S.Y.). We thank Dr T. Matsuzawa, Dr M. Tomonaga, Dr T. Humle and many other colleagues at the Section of Language and Intelligence, the Primate Research Institute (PRI), Kyoto University for their support and helpful comments on the present study. We also thank Dr J. Suzuki, Dr T. Miyabe, Ms A. Watanabe, Mr A. Kaneko, Mr K. Kumazaki, Mr N. Maeda, Mr S. Watanabe, Mr R. Hirokawa, Mr Y. Furuhashi, Ms H. Sasaki and other staff members for veterinary and daily care of the chimpanzees at the PRI.
References
- S.T. Boysen, G.G. Berntson Responses to quantity: perceptual versus cognitive mechanisms in chimpanzees (Pan troglodytes) Journal of Experimental Psychology, 21 (1995), pp. 82–86
- S.T. Boysen, G.G. Berntson, M.B. Hannan, J.T. Cacioppo Quantity-based interference and symbolic representations in chimpanzees (Pan troglodytes) Journal of Experimental Psychology, 22 (1996), pp. 76–86
- S.T. Boysen, K.L. Mukobi, G.G. Berntson Overcoming response bias using symbolic representations of number by chimpanzees (Pan troglodytes) Animal Learning and Behavior, 27 (1999), pp. 229–235
- J. Bräuer, J. Call, M. Tomasello Are apes really inequity averse? Proceedings of the Royal Society B, 273 (2006), pp. 3123–3128
- S.F. Brosnan, M.J. Beran Trading behavior between conspecifics in chimpanzees, Pan troglodytes Journal of Comparative Psychology, 123 (2009), pp. 181–194
- S.F. Brosnan, F.B.M. de Waal A proximate perspective on reciprocal altruism Human Nature, 13 (2002), pp. 129–152
- S.F. Brosnan, H.C. Schiff, F.B.M. de Waal Tolerance for inequity may increase with social closeness in chimpanzees Proceedings of the Royal Society B, 272 (2005), pp. 253–258
- S.F. Brosnan, J.B. Silk, J. Henrich, M.C. Mareno, S.P. Lambeth, S.J. Schapiro Chimpanzees (Pan troglodytes) do not develop contingent reciprocity in an experimental task Animal Cognition, 12 (2009), pp. 587–597
- J.M. Burkart, E. Fehr, C. Efferson, C.P. van Schaik Other-regarding preferences in a non-human primate: common marmosets provision food altruistically Proceedings of the National Academy of Sciences, U.S.A., 104 (2007), pp. 19764–19766
- M.K. Chen, M. Hauser Modeling reciprocation and cooperation in primates: evidence for a punishing strategy Journal of Theoretical Biology, 235 (2005), pp. 5–12
- K.A. Cronin, K.K.E. Schroeder, E.S. Rothwell, J.B. Silk, C.T. Snowdon Cooperatively breeding cottontop tamarins (Saguinus oedipus) do not donate rewards to their long-term mates Journal of Comparative Psychology, 123 (2009), pp. 231–241
- K.G. Duffy, R.W. Wrangham, J.B. Silk Male chimpanzees exchange political support for mating opportunities Current Biology, 17 (2007), pp. R586–R587
- I.C. Gilby Meat sharing among the Gombe chimpanzees: harassment and reciprocal exchange Animal Behaviour, 71 (2006), pp. 953–963
- J. Goodall The Chimpanzees of Gombe: Patterns of Behavior Harvard University Press, Cambridge, Massachusetts (1986)
- P. Hammerstein Why is reciprocity so rare in social animals? A protestant appeal P. Hammerstein (Ed.), Genetic and Cultural Evolution of Cooperation, MIT Press, Cambridge, Massachusetts (2003), pp. 83–93
- W.D. Hamilton The genetical evolution of social behavior. 1 and 2 Journal of Theoretical Biology, 7 (1964), pp. 17–52
- M.D. Hauser, M.K. Chen, F. Chen, E. Chuang Give unto others: genetically unrelated cotton-top tamarin monkeys preferentially give food to those who altruistically give food back Proceedings of the Royal Society, B, 270 (2003), pp. 2363–2370
- K. Hikami, T. Matsuzawa Lever pressing of two chimpanzees in a competitive/cooperative situation Primate Research, 7 (1991), pp. 12–22 (in Japanese with English abstract)
- S. Hirata Chimpanzee social intelligence: selfishness, altruism, and the mother–infant bond Primates, 50 (2009), pp. 3–11
- K. Jensen, B. Hare, J. Call, M. Tomasello What's in it for me? Self-regard precludes altruism and spite in chimpanzees Proceedings of the Royal Society, B, 273 (2006), pp. 1013–1021
- K. Jensen, J. Call, M. Tomasello Chimpanzees are rational maximizers in an ultimatum game Science, 318 (2007), pp. 107–109
- N.F. Koyama, C. Caws, F. Aureli Interchange of grooming and agonistic support in chimpanzees International Journal of Primatology, 27 (2006), pp. 1293–1309
- V.R. Lakshminarayanan, L.R. Santos Capuchin monkeys are sensitive to others' welfare Current Biology, 18 (2008), pp. 999–1000
- K.E. Langergraber, J.C. Mitani, L. Vigilant The limited impact of kinship on cooperation in wild chimpanzees Proceedings of the National Academy of Sciences, U.S.A., 104 (2007), pp. 7786–7790
- T. Matsuzawa The Ai project: historical and ecological contexts Animal Cognition, 6 (2003), pp. 199–211
- T. Matsuzawa The chimpanzee mind: in search of the evolutionary roots of the human mind Animal Cognition, 12 (2009), pp. S1–S9
- T. Matsuzawa, D. Biro, T. Humle, N. Inoue-Nakamura, R. Tonooka, G. Yamakoshi Emergence of culture in wild chimpanzees: education by master-apprenticeship T. Matsuzawa (Ed.), Primate Origins of Human Cognition and Behavior, Springer-Verlag, Tokeyo (2001), pp. 557–574
- A. Melis, B. Hare, M. Tomasello Do chimpanzees reciprocate received favours? Animal Behaviour, 76 (2008), pp. 951–962
- J.C. Mitani, D.P. Watts Why do chimpanzees hunt and share meat? Animal Behaviour, 61 (2001), pp. 915–924
- J.C. Mitani, D.A. Merriwether, C. Zhang Male affiliation, cooperation and kinship in wild chimpanzees Animal Behaviour, 59 (2000), pp. 885–893
- J.B. Silk, S.F. Brosnan, J. Vonk, J. Henrich, D.J. Povinelli, A.S. Richardson, S.P. Lambeth, J. Mascaro, S.J. SchapiroChimpanzees are indifferent to the welfare of unrelated group members Nature, 437 (2005), pp. 1357–1359
- J.R. Stevens The selfish nature of generosity: harassment and food sharing in primates Proceedings of the Royal Society, B, 271 (2004), pp. 451–456
- J.R. Stevens, I.C. Gilby A conceptual framework for nonkin food sharing: timing and currency of benefits Animal Behaviour, 67 (2004), pp. 603–614
- M. Tanaka, S. Yamamoto Token transfer between mother and offspring chimpanzees (Pan troglodytes): mother–offspring interaction in a competitive situation Animal Cognition, 12 (2009), pp. S19–S26
- R.L. Trivers The evolution of reciprocal altruism Quarterly Review of Biology, 46 (1971), pp. 25–57
- A. Ueno, T. Matsuzawa Food transfer between chimpanzee mothers and their infants Primates, 45 (2004), pp. 231–239
- J. Vonk, S.F. Brosnan, J.B. Silk, J. Henrich, A.S. Richardson, S.P. Lambeth, S.J. Schapiro, D.J. Povinelli Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members Animal Behaviour, 75 (2008), pp. 1757–1770
- F.B.M. de Waal Good Natured Harvard University Press, Cambridge, Massachusetts (1996)
- F.B.M. de Waal The chimpanzee's service economy: food for grooming Evolution and Human Behavior, 18 (1997), pp. 375–386
- F.B.M. de Waal Attitudinal reciprocity in food sharing among brown capuchin monkeys Behaviour, 60 (2000), pp. 253–361
- F.B.M. de Waal, M.L. Berger Payment for labour in monkeys Nature, 404 (2000), p. 563
- F.B.M. de Waal, L.M. Luttrell Mechanisms of social reciprocity in three primate species: symmetrical relationship characterisics or cognition? Ethology and Sociobiology, 9 (1988), pp. 101–118
- F.B.M. de Waal, K. Leimgruber, A.R. Greenberg Giving is self-rewarding for monkeys Proceedings of the National Academy of Sciences, U.S.A., 105 (2008), pp. 13685–13689
- F. Warneken, M. Tomasello Altruistic helping in human infants and young chimpanzees Science, 311 (2006), pp. 1301–1303
- F. Warneken, B. Hare, A.P. Melis, D. Hanus, M. Tomasello Spontaneous altruism by chimpanzees and young children PLoS Biology, 5 (2007), p. e184
- S. Yamamoto, M. Tanaka How did altruism and reciprocity evolve in humans? Perspectives from experiments on chimpanzees (Pan troglodytes) Interaction Studies, 10 (2009), pp. 150–182
- S. Yamamoto, M. Tanaka Do chimpanzees (Pan troglodytes) spontaneously take turns in a reciprocal cooperation task? Journal of Comparative Psychology, 123 (2009), pp. 242–249
- S. Yamamoto, M. Tanaka Development of selfish tactics but not reciprocal cooperation in chimpanzee (Pan troglodytes) mother–infant pairs in social problem situations Animal Cognition, 12 (2009), pp. S27–S36
- S. Yamamoto, T. Humle, M. Tanaka Chimpanzees help each other upon request PLoS ONE, 4 (2009), p. e7416